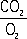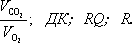Дынное дерево
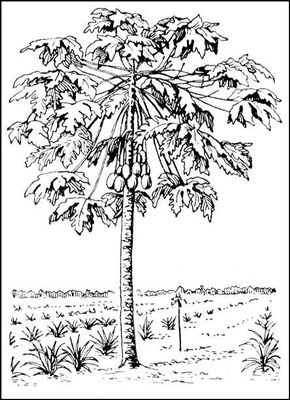
Ды'нное де'рево,папайя (Carica papaya), плодовое дерево семейства папайевых. Листья крупные, пальчаторассечённые, 5—7-лопастные на длинных черешках, расположены пучками на верхушке неветвящегося невысокого (4—8
м) ствола. Цветки желтовато-белые, раздельнополые (тычиночные — в кистях, пестичные — большей частью одиночные, пазушные), обычно двудомные или обоеполые. Плоды напоминают дыню. Д. д. быстро растёт, недолговечно, неприхотливо к почвам, не выносит морозов. В диком состоянии неизвестно. Разводится в тропиках. Плоды Д. д. используют в пищу. Из их млечного сока получают фермент
.
Лит.:Синягин И. И., Тропическое земледелие, М., 1968.
С. К. Черепанов.
Плод дынного дерева.
Дынное дерево.
Дынька
Ды'нька(в архитектуре), декоративная деталь (утолщение) столбов, колонок, наличников окон и дверных порталов. Д. использовались главным образом в русской архитектуре 15—17 вв. В деревянном зодчестве Д. применялись до конца 18 в.
Дыньки портала в церкви Иоанна Златоуста в Коровниках в Ярославле (1649—54).
Дыня
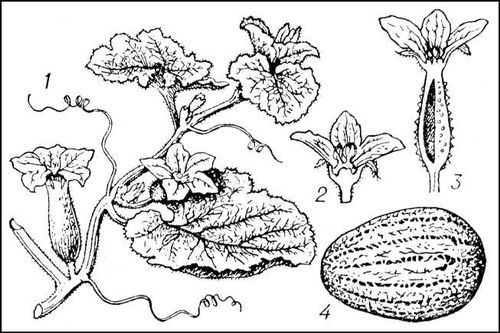
Ды'ня,однолетнее травянистое растение рода Cucumis семейства тыквенных. Стебель округло-гранёный. Листья крупные на длинных черешках, в пазухах листьев усики. Цветки раздельнополые, но встречаются и обоеполые. Опыление с помощью насекомых. Плод — многосемянная тыквина разнообразной формы (от 200
гдо 16
кгв зависимости от сорта). Д. происходит из Средней и Малой Азии. Известно несколько видов, по различным классификациям от 1 до 15. Наиболее распространена в культуре Д. обыкновенная, или столовая (С. melo). Теплолюбивое растение. Рост и развитие нормально происходят при температуре воздуха 25—30°С, при снижении до 15°С развитие замедляется, а при 3—5°С растения погибают. Д. засухоустойчива. Культивируется в США, Индии, Иране, Афганистане, Малой Азии, Японии, на Балканах, в Испании, Италии, Франции и др. В СССР Д. разводят в республиках Средней Азии, Поволжье, Молдавии, на Украине и в Закавказье. Плоды Д. столовой — ценный пищевой и диетический продукт, содержат сахар (16—18% и более), витамин С (до 60
мг%), каротин, пектиновые вещества, минеральные соли. Плоды употребляют в свежем виде, вялят, сушат; используют также для приготовления варенья, цукатов, мармелада, повидла, бекмеса (дынного мёда). Урожай плодов в орошаемых условиях (Средняя Азия) 300—400
цс 1
гаи выше, в неорошаемых (Поволжье, Украина и др.) до 200
цс 1
га.
Сорта Д. столовой разделяют на группы: аданы, хандаляки, амери, канталупы, кассабы, зарды. В республиках Средней Азии и Казахстане возделывают: ранние сорта — Хандаляк жёлтая местная, Босвалды, Бухарка 944, Заами 672 и др.; летние — Арбакешка 1219, Амери 696, Кокча 588; осенние — Гуляби оранжевая, Кара-пучак 3744, Кой-баш 476; зимние — Гуляби зелёная, Гуляби-кара 694, Гуляби-сары 497 и многие др. В Европейской части СССР наиболее распространены сорта: ранние — Лимонно-жёлтая, Новинка Кубани; среднего и среднепозднего созревания — Колхозница 749/753, Колхозница 593, Быковская 735, Украинка; позднего и среднепозднего созревания — Зимовка с яблочными семенами и др. В Закавказье выращивают в основном местные сорта: раннеспелые — Салвар местная, Мегрули местная и др.; среднеспелые — Снейваз местная, Масис 2 и др.
Под Д. отводят темноцветные супеси и лёгкие суглинки — в засушливой зоне Нижнего Поволжья, лёгкие или средние суглинки и супесчаные чернозёмы на Северном Кавказе и в степной зоне Украины, супесчаные удобренные почвы — в районах Средней Азии. Перед посевом семена Д. прогревают на солнце 6—7 дней или искусственно при температуре 50—60°С в течение 4—6
ч. Посев квадратно-гнездовой или рядовой, площадь питания 3—4
м
2на неполивных землях и 2—3
м
2на поливных. Семена высевают, когда почва прогреется до температуры 12—14°С. В северных районах бахчеводства применяют рассадную культуру Д. Рассаду выращивают в торфоперегнойных горшках в парниках и после прекращения весенних заморозков высаживают в открытый грунт. На посевах проводят 1—2 прорывки растений, до трёх междурядных обработок почвы, 1—2 подкормки растений, в основном фосфорными удобрениями, расправку и присыпку плетей для предохранения их от переворачивания ветром, а на орошаемых бахчах, кроме того, рыхление борозд после каждого полива. Убирают плоды выборочно, по мере созревания, а у позднеспелых сортов — недозрелые, которые поспевают при хранении.
Лит.:Дудко П. Н., Сортовое богатство дынь Узбекистана, Таш., 1956; Пангало К. Т., Дыни, Киш., 1958; Юрина О. В., Селекция и семеноводство тыквенных культур, М., 1966.
В. Ф. Белик.
Дыня: 1 — общий вид растения; 2 — тычиночный цветок; 3 — пестичный цветок; 4 — плод.
Дыравка
Дыра'вка,род травянистых растений семейства мальвовых, известных в садоводстве под названием
.
Дырка
Ды'рка(в зонной теории
), не занятое электроном энергетическое состояние, например в валентной зоне
. Д. ведут себя как частицы (
) с положительным зарядом, равным по абсолютной величине заряду электрона, и являются наряду с электронами носителями тока в полупроводниках (дырочная электропроводность).
Лит. см. при ст.
,
.
Дыхание
Дыха'ние,совокупность процессов, которые обеспечивают поступление в организм кислорода и выделение из него углекислого газа (внешнее Д.) и использование кислорода клетками и тканями для окисления органических веществ с освобождением содержащейся в них энергии, необходимой для жизнедеятельности (
, клеточное Д.). Бескислородный путь освобождения энергии свойствен только небольшой группе организмов — так называемым
(см.
); в ходе эволюции освобождение энергии в результате Д. стало у подавляющего большинства организмов главным процессом, а анаэробные реакции сохранились в основном как промежуточные этапы
.
Д. животных и человека.У простейших, губок, кишечнополостных и некоторых др. организмов О
2диффундирует непосредственно через поверхность тела. С усложнением организации и увеличением размеров тела появляются специальные
, а также система кровообращения, в которой циркулирует жидкость — кровь или гемолимфа, содержащая вещества, способные связывать и переносить O
2и CO
2(см.
). У насекомых O
2поступает в ткани из системы воздухоносных трубочек —
. У водных животных, использующих растворённый в воде O
2, органами Д. служат
, снабжённые богатой сетью кровеносных сосудов. В этом случае O
2, растворённый в воде, диффундирует в кровь, циркулирующую в сосудах жаберных щелей. У многих рыб значительную роль играет кишечное Д., при котором воздух заглатывается и O
2поступает в кровь через кровеносные сосуды кишечника; некоторую роль в Д. рыб играет также плавательный пузырь; у многих обитающих в воде животных обмен газов (главным образом СО
2) происходит и через кожу. У сухопутных животных внешнее Д. обеспечивается преимущественно
. У земноводных и многих др. животных наряду с этим функционирует кожное Д. У птиц существенное значение имеют сообщающиеся с лёгкими
, которые изменяются в объёме при летательных движениях и облегчают Д. в полёте. У земноводных и пресмыкающихся воздух в лёгкие нагнетается движениями мышц дна рта. У птиц, млекопитающих и человека внешнее Д. обеспечивается ритмической работой дыхательных мышц (главным образом диафрагмы и межрёберных мышц), координируемой нервной системой. При сокращении этих мышц объём грудной клетки увеличивается и происходит растяжение находящихся в ней лёгких; поэтому возникает разность между атмосферным и внутрилёгочным давлением и воздух поступает в лёгкие (вдох). Выдох может быть пассивным — за счёт спадения растянутой во время вдоха грудной клетки, а вслед за ней и лёгких; активный выдох обусловлен сокращением некоторых групп мышц. Количество воздуха, поступающее в легкие за 1 вдох, называется дыхательным объёмом (см.
).
При Д. дыхательная мускулатура преодолевает эластичное сопротивление, связанное с упругостью грудной клетки, тягой лёгких и поверхностным натяжением альвеол. Последнее, однако, значительно снижается под влиянием поверхностно активного вещества, вырабатываемого клетками альвеолярного эпителия; поэтому альвеолы при выдохе не спадаются, а при вдохе легко расширяются. Чем выше эластичное сопротивление, тем труднее растягиваются грудная клетка и лёгкие; при глубоком Д. работа дыхательной мускулатуры, затрачиваемая на его преодоление, резко возрастает. Неэластичное сопротивление Д. обусловлено главным образом трением воздуха при его движении по носовым ходам, гортани, трахее и бронхам. Оно зависит от скорости потока воздуха во время Д. и от его характера. При спокойном Д. поток близок к ламинарному (линейному) в прямых участках воздухоносных путей и к турбулентному (вихревому) в местах разветвления или сужения. С увеличением скорости потока (при форсированном Д.) турбулентность возрастает и для продвижения воздуха требуется более высокая разность давлений, а следовательно, и увеличение работы дыхательных мышц. Неравномерное распределение сопротивления движению воздуха по дыхательным путям приводит к тому, что поступление воздуха в разные группы лёгочных альвеол происходит неравномерно; такая разница в вентиляции особенно значительна при лёгочных заболеваниях.
Количество воздуха, вентилирующее лёгкие в 1
мин, называется минутным объёмом дыхания (МОД). МОД равен произведению дыхательного объёма на частоту Д. (число дыхательных движений в 1
мин, равное у человека примерно 15—18) и составляет у взрослого человека в покое 5—8
л/мин. Только часть МОД (около 70%) участвует в обмене газов между вдыхаемым и альвеолярным воздухом, эту часть называют объёмом альвеолярной вентиляции; остальная часть МОД используется для «промывания» так называемого мёртвого, или вредного, пространства дыхательных путей, в котором к началу выдоха сохраняется наружный воздух, заполнивший его в конце предшествовавшего вдоха (объём мёртвого пространства около 160
мл). Вентиляция альвеол обеспечивает постоянство состава
. Парциальное давление O
2(
p
O2) и CO
2(
p
CO2) в альвеолярном воздухе колеблется в очень узких пределах и составляет для О
2около 13
кн/м
2(100
мм рт. cт.) и для СО
2около 5,4
кн/м
2(40
мм рт. ст.).
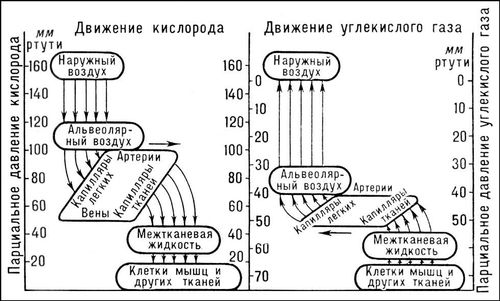
Обмен газов между альвеолярным воздухом и венозной кровью, поступающей в капилляры лёгких, осуществляется через альвеоло-капиллярную мембрану, общая поверхность которой очень велика (у человека около 90
м
2). Диффузия O
2в кровь обеспечивается разностью парциальных давлений O
2в альвеолярном воздухе и в венозной крови (8—9
кн/м
2, или 60—70
мм рт. ст.). CO
2, приносимый кровью из тканей в связанной форме (бикарбонаты, соли угольной кислоты и карбгемоглобин), освобождается в капиллярах лёгких при участии фермента карбоангидразы и диффундирует из крови в альвеолы; разность
p
CO2между венозной кровью и альвеолярным воздухом составляет около 7
мм рт. ст.Способность альвеолярной стенки пропускать O
2и CO
2, так называемая диффузионная способность лёгких, очень велика: в покое она составляет в 1
минпримерно 30
млO
2на 1
ммразности
p
O2между альвеолярным воздухом и кровью; для CO
2эта величина во много раз больше. Поэтому парциальное давление газов в оттекающей из лёгких артериальной крови успевает приблизиться к их давлению в альвеолярном воздухе. Переход O
2в ткани и удаление из них CO
2также происходят путём диффузии, т.к.
p
O2в тканевой жидкости 2,7—5,4
кн/м
2(20—40
мм рт. ст.), а в клетках ещё ниже, а
p
CO2в клетках может достигать 60
мм рт. cт.(см.
рис.
).
Потребление O
2клетками и тканями и образование ими CO
2, что составляет сущность тканевого, или клеточного, Д., — одна из основных форм
, осуществляющейся у животных и растений в принципе одинаково. Высокое потребление O
2характерно для тканей почек, коры больших полушарий головного мозга, сердца. В результате
тканевого Д. освобождается энергия, расходуемая на все жизненные проявления. Процесс этот осуществляется в
и складывается из дегидрирования субстратов Д. — углеводов и продуктов их расщепления, жиров и жирных кислот, аминокислот и продуктов их дезаминирования. Субстраты Д. поглощают O
2и служат источником CO
2(отношение —
называется
). Энергия, освобождающаяся при окислении органических веществ, не используется тканями непосредственно, т.к. около 70% её расходуется на образование АТФ — одной из
, последующее ферментативное расщепление которой обеспечивает энергетические потребности тканей, органов и организма в целом (см.
,
). Т. о., с биохимической точки зрения Д. — это превращение энергии углеводов и др. веществ в энергию макроэргических фосфатных связей.
Постоянство
p
O2и
p
CO2в альвеолярном воздухе, а стало быть, и в артериальной крови может поддерживаться лишь при условии, если альвеолярная вентиляция соответствует скорости потребления организмом O
2и образования CO
2, т. е. уровню обмена веществ. Это условие обеспечивается благодаря совершенным механизмам регуляции Д. Управление частотой и глубиной Д. осуществляется рефлекторным путём. Так, повышение
p
CO2и снижение
p
O2в альвеолярном воздухе и в артериальной крови возбуждают хеморецепторы синокаротидной и кардиоаортальной зон, что приводит к возбуждению
и увеличению МОД. Согласно классическим представлениям, повышение
p
CO2в артериальной крови, омывающей дыхательный центр, также возбуждает его и вызывает увеличение МОД. Т. о., регуляция Д. по отклонению
p
O2и
p
CO2в артериальной крови, осуществляемая по типу обратной связи, обеспечивает оптимальный МОД. Однако в ряде случаев, например при мышечной работе, МОД увеличивается до наступления в обмене веществ сдвигов, которые приводят к изменениям в газовом составе крови. Это усиление вентиляции обусловлено сигналами, поступающими в дыхательный центр от рецепторов двигательного аппарата, двигательной зоны коры больших полушарий мозга, а также
на различные сигналы, связанные с привычной работой и её обстановкой. Т. о., управление Д. осуществляется сложной самообучающейся системой не только по принципу регулирования по отклонению, но и по сигналам, предупреждающим о возможных отклонениях. Смена вдоха и выдоха обеспечивается системой взаимодополняющих механизмов. Во время вдоха в дыхательный центр по волокнам блуждающих нервов поступают импульсы от рецепторов растяжения, находящихся в лёгких. При достижении лёгкими определённого объёма эта импульсация тормозит клетки дыхательного центра, возбуждение которых вызывает вдох. При выключении нервных путей, обеспечивающих поступление импульсов в дыхательный центр, ритмичность Д. сохраняется благодаря автоматизму центра, однако характер ритма резко отличается от нормального. При нарушениях Д. и механизмов его регуляции возникают изменения газового состава крови (см.
).
Методы исследования Д. разнообразны. В физиологии труда и спорта, клинической медицине широко применяют регистрацию глубины и частоты дыхательных движений, измерения газового состава выдыхаемого воздуха, артериальной крови, плеврального и альвеолярного давления. См. также
.
Лит.:Сеченов И. М., Избр. труды, М., 1935; Холден Дж. и Пристли Дж., Дыхание, пер. с англ., М.—Л., 1937; Маршак М. Е., Регуляция дыхания у человека, М., 1961; Физиология человека, М., 1966; Comroe J. Н., Physiology of respiration, Chi., 1966; Dejours P., Respiration, Oxf., 1966.
Л. Л. Шик.
Д. растений.Д. присуще всем органам, тканям и клеткам растения. Об интенсивности Д. можно судить, измеряя количество выделяемого тканью CO
2либо поглощаемого ею O
2. Более интенсивно дышат молодые, быстро растущие органы и ткани растений. Наиболее активно Д.
, затем листьев; слабее Д. стеблей и корней. Теневыносливые растения дышат слабее светолюбивых. Для высокогорных растений, адаптированных к пониженному парциальному давлению O
2, характерна повышенная интенсивность Д. Очень активно Д. плесневых грибов, бактерий. Д. усиливается с повышением температуры (на каждые 10°С — примерно в 2—3 раза), прекращаясь при 45—50°С. В тканях зимующих органов растений (почки лиственных деревьев, иглы хвойных) Д. продолжается (с резко сниженной интенсивностью) и при значительных морозах. Д. стимулируют механические и химические раздражения (поранения, некоторые яды, наркотики и т.п.). Закономерно изменяется Д. в ходе развития растения и его органов. Сухие (покоящиеся) семена дышат очень слабо; при набухании и последующем прорастании семян Д. усиливается в сотни и тысячи раз. С окончанием периода активного роста растений Д. их тканей ослабевает, что связано с процессом старения протоплазмы. При созревании семян, плодов интенсивность Д. уменьшается.
Согласно теории советского биохимика А. Н. Баха, процесс Д., т. е. окисление углеводов, жиров, белков, осуществляется при помощи окислительной системы клетки в два этапа: 1) активирование O
2воздуха путём его присоединения к содержащимся в живой клетке ненасыщенным, способным самопроизвольно окисляться соединениям (оксигеназам) с образованием перекисей; 2) активирование последних с освобождением атомарного кислорода, способного окислять трудно окисляемые органические вещества. По теории дегидрирования русского ботаника В. И. Палладина, важнейшее звено Д. — активация водорода субстрата, осуществляемая
. Обязательный участник сложной цепи процессов Д. — вода, водород которой вместе с водородом субстрата используется для восстановления самоокисляющихся соединений — так называемых дыхательных пигментов. CO
2, выделяющийся при Д., образуется без участия кислорода воздуха, т. е. анаэробно. Кислород воздуха идёт на окисление дыхательных хромогенов, превращающихся при этом в дыхательные пигменты. Дальнейшее развитие теория Д. получила в исследованиях советского ботаника С. П.
, согласно которым первые этапы аэробного Д. аналогичны процессам, свойственным анаэробам. Превращения образующегося при этом промежуточного продукта могут идти с участием кислорода, что свойственно аэробам. У анаэробов же эти превращения идут без участия молекулярного кислорода. По современным представлениям, процесс окисления, который составляет химическую основу Д., заключается в потере веществом электрона. Способность присоединять или отдавать электроны зависит от величины окислительного потенциала соединения. Кислород обладает самым высоким окислительным потенциалом и, следовательно, максимальной способностью присоединять электроны. Однако потенциал кислорода сильно отличается от потенциала дыхательного субстрата. Поэтому роль промежуточных переносчиков электронов от дыхательного субстрата к кислороду выполняют специфические соединения. Попеременно окисляясь и восстанавливаясь, они образуют систему переноса электронов. Присоединив к себе электрон от менее окисленного компонента, такой переносчик восстанавливается и, отдавая его следующему компоненту с более высоким потенциалом, окисляется. Так электрон передаётся от одного звена дыхательной цепи к другому и, в конце концов, кислороду. Таков заключительный этап Д.
Все эти процессы (активация кислорода, водорода, перенос электрона по цепи на кислород) осуществляются главным образом в митохондриях благодаря разветвлённой системе окислительно-восстановительных ферментов (см.
). По пути следования к кислороду электроны, мобилизуемые первоначально от молекулы органического вещества, постепенно отдают заключённую в них энергию, которую клетка запасает в форме химических соединений, главным образом АТФ.
Благодаря совершенным механизмам запасания и использования энергии процессы энергообмена в клетке идут с очень высоким кпд, пока недостижимым в технике. Биологическая роль Д. не исчерпывается использованием энергии, заключённой в окисляемой органической молекуле. В ходе окислительных превращений органических веществ образуются активные промежуточные соединения — метаболиты, которые живая клетка использует для синтеза специфических составных частей своей протоплазмы, образования ферментов и др. Всем этим определяется центральное место, занимаемое Д. в комплексе процессов обмена веществ живой клетки. В Д. скрещиваются и увязываются процессы обмена белков, нуклеиновых кислот, углеводов, жиров и др. компонентов протоплазмы.
Лит.:Костычев С. П., Физиология растений, 3 изд., т. 1, М.—Л., 1937; Бах А. Н., Собр. трудов по химии и биохимии, М., 1950; Таусон В. О., Основные положения растительной биоэнергетики, М.—Л., 1950; Джеймс В. О., Дыхание растений, пер. с англ., М., 1956; Палладин В. И., Избр. труды, М., 1960; Михлин Д. М., Биохимия клеточного дыхания, М., 1960; Сент-Дьердьи А., Биоэнергетика, пер. с англ., М., 1960; Рубин Б. А., Ладыгина М. Е., Энзимология и биология дыхания растений, М., 1966; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Рубин Б. А., Курс физиологии растений, 3 изд., М., 1971; Кретович В. Л., Основы биохимии растений, М., 1971.
Б. А. Рубин.
Рис. к ст. Дыхание.
Дыхания органы
Дыха'ния о'рганы,органы, служащие для
между организмом и внешней средой. Д. о. имеются только у животных —
, получающих свободный кислород или непосредственно из атмосферного воздуха (воздушное дыхание), или из воздуха, растворённого в воде (водное дыхание). У анаэробных организмов (одно- и многоклеточных) Д. о. отсутствуют. Многие группы низших аэробных организмов также не имеют Д. о. (простейшие, губки, кишечнополостные, многие черви). У животных с тонкими нежными покровами кислород может проникать в организм путём диффузии через всю поверхность тела — кожный тип дыхания, к которому часто присоединяется кишечное дыхание, когда кислород поступает в организм через стенку кишечного канала (например, у кишечнополостных). Среди позвоночных кожное дыхание играет большую роль у многих рыб и земноводных. Происхождение и строение Д. о. у разных организмов различны. Д. о. воздушного типа называются
и
, Д. о. водного типа —
.
Особый интерес представляют имеющиеся у рыб, живущих в иле (некоторых Clupeidae, Characinidae, Siluridae), дополнительные Д. о. — гроздевидные выросты, отходящие от жаберных дужек и служащие для воздушного дыхания. У лабиринтовых рыб на первой жаберной дужке развиваются костные пластинки, покрытые слизистой оболочкой, через которую поглощается O
2. Это приспособление даёт возможность рыбе-ползуну (Anabas) долгое время находиться на суше.
У многих рыб из верхней части жаберных мешков могут вырастать воздушные пузыри, напоминающие настоящее лёгкое (у Amphipnous, Saccobranchus и др.). В процессе индивидуального развития у многих форм происходит смена различных Д. о. Так, среди насекомых у развивающихся в воде подёнок сначала имеет место диффузное кожное дыхание, заменяющееся впоследствии дыханием при помощи трахейных жабр, и, наконец, появляется свойственное взрослому насекомому трахейное воздушное дыхание. У личинок рыб до окончательного развития жабр дыхательную функцию выполняет вначале желточный мешок, снабжённый богатой сетью кровеносных сосудов, затем — сосуды непарных плавников, а иногда временные (личиночные) наружные жабры. У земноводных в личиночной стадии для дыхания служит сеть кровеносных сосудов желточного мешка, в дальнейшем обычно развиваются наружные жабры и часто возникают сосудистые сети в непарном (хвостовом) плавнике. Все зародыши
сначала дышат при помощи кровеносной сети желточного мешка. Впоследствии к такому дыханию присоединяется и вытесняет его так называемое аллантоидное дыхание, когда в стенках
развивается густая сеть сосудов. Только после вылупления из яйца (пресмыкающиеся, птицы) или рождения (млекопитающие животные и человек) начинают функционировать их окончательные Д. о. — лёгкие (см.
).
А. Н. Дружинин.
Дыхательное горло
Дыха'тельное го'рло,начальный отдел нижних дыхательных путей, то же, что
.
Дыхательные мышцы
Дыха'тельные мы'шцы,скелетные мышцы, увеличивающие и уменьшающие объём грудной клетки при дыхании. Основная работа Д. м. приходится на процесс вдоха, т.к. выдох происходит пассивно, вследствие тяжести стенок грудной клетки, эластичности лёгких и межрёберных мышц (в форсированном выдохе участвуют внутренние межрёберные мышцы и мышцы брюшного пресса). Основной мышцей, осуществляющей вдох, является диафрагма; в акте вдоха принимают участие также наружные межрёберные мышцы. Усиленному вдоху способствуют некоторые мышцы шеи и поверхностные мышцы груди. В акт форсированного дыхания вовлекаются также некоторые мышцы спины и мышцы плечевого пояса.
Дыхательные пигменты
Дыха'тельные пигме'нты,группа соединений растительной клетки, участвующих в процессе
; см.
.
Дыхательные хромогены
Дыха'тельные хромоге'ны,группа соединений растительной клетки, способных обратимо окисляться, превращаясь в дыхательные пигменты, которые, восстанавливаясь, вновь превращаются в хромогены. Согласно теории В. И. Палладина, Д. х. и дыхательные пигменты образуют единую систему и выполняют в процессе дыхания роль переносчиков водорода на кислород.
Дыхательный коэффициент
Дыха'тельный коэффицие'нт,отношение объёма выделяемого из организма углекислого газа к объёму поглощаемого за то же время кислорода. Обозначается:
Определение ДК важно для исследования особенностей
и обмена веществ у животных и растительных организмов. При окислении в организме углеводов и полном доступе кислорода ДК равен 1, жиров — 0,7, белков — 0,8. У здорового человека в покое ДК равен 0,85 ± 0,1; при умеренной работе, а также у животных, питающихся преимущественно растительной пищей, приближается к 1. У человека при очень длительной работе, голодании, у плотоядных животных (хищников), а также при спячке, когда из-за ограниченности запасов углеводов в организме усиливается диссимиляция жиров, ДК составляет около 0,7. ДК превышает 1 при интенсивном отложении в организме жиров, образующихся из поступающих с пищей углеводов (например, у человека при восстановлении нормального веса после голодания, после длительных заболеваний, а также у животных при откорме). До 2 ДК возрастает при усиленной работе и гипервентиляции лёгких, когда из организма выделяется дополнительно СО
2, находившийся в связанном состоянии. Ещё больших величин ДК достигает у
, у которых большая часть выделяемого CO
2образуется путём бескислородного окисления (брожения). ДК ниже 0,7 бывает при заболеваниях, связанных с нарушениями обмена веществ, после тяжёлой физической работы.
Л. Л. Шик.
У растений ДК зависит от химической природы дыхательного субстрата, содержания CO
2и O
2в атмосфере и др. факторов, характеризуя, т. о., специфику и условия
. При использовании клеткой для дыхания углеводов (проростки злаков) ДК равен примерно 1, жиров и белков (прорастающие семена масличных и бобовых) — 0,4—0,7. При недостатке О
2и затруднённом его доступе (семена с твёрдой оболочкой) ДК равен 2—3 и более; высокий ДК характерен также для клеток точек роста.
Б. А. Рубин.
Дыхательный центр
Дыха'тельный центр,совокупность нескольких групп нервных клеток (нейронов), расположенных в разных отделах центральной нервной системы, преимущественно в
продолговатого мозга. Постоянная координированная ритмическая активность этих нейронов обеспечивает возникновение дыхательных движений и их регуляцию в соответствии с возникающими в организме изменениями. Импульсы от Д. ц. поступают в двигательные нейроны передних рогов шейного и грудного отделов спинного мозга, от которых возбуждение передаётся к дыхательной мускулатуре. Активность Д. ц. регулируется гуморально, т. е. составом омывающей его крови и тканевой жидкости, и рефлекторно, в ответ на импульсы, поступающие от рецепторов в дыхательной, сердечно-сосудистой, двигательной и др. системах, а также от высших отделов центральной нервной системы. При прекращении поступления нервных импульсов Д. ц. сохраняет активность (так называемый
Д. ц.), способную обеспечить дыхательные движения. См.
.
Дыхсу
Дыхсу',долинный ледник на северном склоне Главного Кавказского хребта, в районе вершин Шхара, Айлама, Фытнаргин. Спускается несколькими потоками до высоты 2040
м. Длина 13
км, площадь 40,7
км
2. Даёт начало р. Дыхсу — одному из истоков р. Черек-Балкарский.
Дыхтау
Дыхта'у,одна из самых высоких вершин в Центральной части Большого Кавказа в Кабардино-Балкарской АССР, близ границы с Грузинской ССР. Находится в Боковом хребте, в истоках р. Черек-Хуламский (Черек-Безенгийский) бассейна Терека. Высота 5203
м. Сложена кристаллическими породами. Альпинизм.