 |
|
���������� ������:: ��� :: ������ ������ ����� :: ������ ������� �������� :: �������� ������� :: ������ ������� :: ������� ��� ���������� :: ������ ����� :: ������� ������ :: �������� ����� :: ������� ������ ���������� �����:: ���������� �� ������� Windows XP :: ������ ���� :: �� ���� ����� �� �����... :: ������, ������, ���� :: ������ � �������� :: ������ ���������� :: ����������� ������ � ������ ������������ :: ���������� c���� :: ��������� ����� � �����-���������� :: ���������� ������ |
������� ��������� ������������ (��)ModernLib.Net / ������������ / ��� / ������� ��������� ������������ (��) - ������ (���. 11)
 и и
 , в мейозе у гетерозиготной формы ® , в мейозе у гетерозиготной формы ®
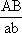 могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами
 и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или
,была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток - с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения
.В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и
совпадает. и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или
,была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток - с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения
.В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и
совпадает.
Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. влияния рентгеновских лучей на возникновение наследственных изменений ( ) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928-30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. В 1957 эти представления были доказаны работой американского учёного С. Бензера с бактериофагом Т4. Использование рентгеновских лучей для стимулирования позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 (открытый в 1925 Стёртевантом), т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы. Х. т. н. развивается в направлении углубления знаний об универсальных носителях наследственной информации - молекулах (ДНК). Установлено, что непрерывная последовательность пуриновых и пиримидиновых оснований вдоль цепи ДНК образует гены, межгенные интервалы, знаки начала и конца считывания информации в пределах гена; определяет наследственный характер синтеза специфических белков клетки и, следовательно, наследственный характер обмена веществ. ДНК составляет материальную основу группы сцепления у бактерий и многих вирусов (у некоторых вирусов носителем наследственной информации является ) ;молекулы ДНК, входящие в состав , и др. клетки, служат материальными носителями цитоплазматической наследственности. Х. т. н., объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в с.-х. науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения Х. т. н. позволяют более рационально вести с.-х. производство. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки - отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование .На знании закономерностей хромосомных перестроек основывается изучение человека. Лит.:Морган Т. Г., Структурные основы наследственности, пер. с англ., М.-П., 1924; его же, Избранные работы по генетике, пер, с англ., М.-Л., 1937; Актуальные вопросы современной генетики, М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Классики советской генетики. [Сб. ст.], Л., 1968. С. Г. Инге-Вечтомов. 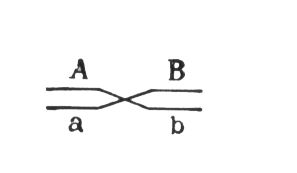
Рис. к ст. Хромосомная теория наследственности. Хромосомные болезни Хромосо'мные боле'зни,наследственные заболевания, обусловленные изменением числа или структуры .Частота Х. б. среди новорождённых детей около 1%. Многие изменения хромосом несовместимы с жизнью и являются частой причиной спонтанных абортов и мертворождений. При спонтанных абортах обнаружено около 20% эмбрионов с аномальными кариотипами (хромосомными наборами). Изменение числа хромосом происходит в результате нерасхождения их в или при делении клеток на ранней стадии развития оплодотворённого яйца (см. ) .Нерасхождению хромосом при первых делениях оплодотворённого яйца способствует, например, высокий возраст матери. Хромосомные обусловливаются физическими (ионизирующее излучение) и химическими (например, лекарственные препараты с мутагенным эффектом) факторами; вирусами (краснухи, вирусного гепатита, ветряной оспы и др.), антителами и различными расстройствами метаболизма. Х. б. могут быть связаны с излишком генетического материала (полисемия - наличие одной или нескольких добавочных хромосом; ; ) ;с утратой части генетического материала ( , , ) ;с хромосомными перестройками ( ;различные перестановки участков хромосом). Различают также группы Х. б., обусловленных изменениями половых и неполовых хромосом. Наиболее распространённые аномалии первой группы у женщин - синдром Шерешевского - Тернера (моносомия Х) и синдром трисомии Х; у мужчин - синдром Клайнфельтера, характеризующийся наличием лишней Х-хромосомы. При синдромах Шерешевского - Тернера и Клайнфельтера возникают задержка полового развития и бесплодие; при синдроме трисомии Х - некоторое снижение интеллекта, расстройства менструального цикла. Частота аномалий по половым хромосомам у мертворождённых составляет 2,7%, что в 25 раз выше, чем среди новорождённых. Среди аутосомных аномалий с нарушением числа хромосом выделяются трисомные синдромы: синдром трисомии хромосом группы D (13-15-е пары), или синдром Патау, встречающийся с частотой 1: 4000 новорождённых; синдром трисомии хромосом группы Е (18-я пара) - Эдвардса, с частотой 1: 300 и (трисомия по 21-й хромосоме), частота которой 1: 700 новорождённых. Указанные Х. б. проявляются различными уродствами; задержкой физического и умственного развития; пороками развития внутренних органов. Отмечается специфическое сочетание отдельных аномалий в различных случаях трисомии. Подобные больные живут, как правило, недолго, погибают от вторичных инфекций. Тяжесть клинической картины при синдромах, вызванных структурными изменениями хромосом, как правило, коррелирует с количеством избыточного или недостающего хромосомного материала. Специфика патологических проявлений зависит от того, какая хромосома вовлечена в процесс перестройки. Чаще отмечаются задержка умственного и физического развития, мышечная гипотония, аномалии лицевого скелета. пороки развития внутренних органов. Наряду с типичными Х. б. описано большое количество (около 200) синдромов, вызванных сложными типами хромосомных аберраций. Единственно надёжный метод диагностики Х. б. - цитогенетическое исследование кариотипа, а при изменении числа половых хромосом - дополнительно исследование .Лечение Х. б. сводится к назначению общеукрепляющих, стимулирующих и поддерживающих средств, в том числе гормонов, и др. В профилактике важную роль играет ,которая позволяет выявить семьи с повышенным риском рождения больного ребёнка. Перспективный метод внутриутробной диагностики хромосомного набора плода повышает эффективность медико-генетической консультации в случаях прогнозирования исхода беременности в семьях с повышенным риском рождения ребёнка, больного Х. б. Лит.см. при статьях , . Хромосомные перестройки Хромосо'мные перестро'йки,хромосомные мутации, структурные изменения хромосом, возникающие вследствие разрывов в них, сопровождающихся в большинстве случаев воссоединением образовавшихся кусков в иных сочетаниях, чем в исходных хромосомах. Структурные перестройки могут происходить как в пределах одной хромосомы, так и между гомологическими и негомологическими хромосомами и состоят в выпадении ( ) или удвоении ( ) какого-либо участка хромосомы, в перенесении его в др. сегмент хромосомы ( ) ;наконец, участок может оказаться перевёрнутым на 180°, оставаясь в той же хромосоме ( ) .Делеции и дупликации нарушают генный баланс, что ведёт к изменению признаков организма. Инверсии, изменяющие лишь порядок расположения генов в хромосоме, и транслокации, при которых гены перемещенного участка попадают в др. группу сцепления, не нарушают генного баланса и не изменяют фенотипических признаков организма. При у ,содержащих одну нормальную хромосому и одну с инверсией, сближение этих хромосом затруднено, между ними подавлен или идёт с пониженной частотой, нередко возникают анеуплоидные гаметы (см. ) ,поэтому такие организмы отличаются пониженной плодовитостью по сравнению с гомозиготами, у которых обе хромосомы данной пары несут инверсию или обе нормальны. Гетерозиготы по хромосомам, несущим транслокацию, дают много анеуплоидных гамет, поэтому плодовитость их ниже, чем у гомозиготных организмов. Х. п. могут возникать спонтанно, но частота их резко возрастает под влиянием физических и химических факторов (см. , ) .Небольшие делеции и дупликации могут быть следствием неравного кроссинговера. Х. п. играют большую роль в эволюции организмов: дупликации представляют главный источник увеличения числа генов; инверсии и транслокации могут вести к генетической изоляции гомозиготных по ним особей, более плодовитых, чем гетерозиготы. При всех Х. п. иногда наблюдается т. н. ,заключающийся в том, что ген, перенесённый в новое место хромосомы, изменяет своё действие на фенотип организма. Х. п. могут быть использованы в практических целях для изменения групп ,определяющих хозяйственно ценные признаки организмов. С. М. Гершензон. Хромосомный набор Хромосо'мный набо'р,совокупность ,заключённая в ядре любой клетки тела растительного или животного организма; характеризуется постоянным для каждого биологического вида числом хромосом, определённой их величиной и морфологическими особенностями. См. . Хромосомы Хромосо'мы(от и ) ,органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор Х. в клетке, характерный для данного организма, называется .В любой клетке тела большинства животных и растений каждая Х. представлена дважды: одна из них получена от отца, другая - от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие Х. называются гомологичными, набор гомологичных Х. - диплоидным. В хромосомном наборе клеток раздельнополых организмов присутствует пара (или несколько пар) половых хромосом, как правило, различающихся у разных полов по морфологическим признакам; остальные Х. называются аутосомами. У млекопитающих в половых Х. локализованы ,определяющие пол организма; у плодовой мушки пол определяется соотношением половых хромосом и аутосом (балансовая теория определения пола). Первоначально Х. были описаны как интенсивно окрашивающиеся основными красителями плотные тельца (немецкий учёный В. Вальдейер, 1888). Однако оказалось, что внешний вид Х. существенно меняется на разных стадиях клеточного цикла, и как компактные образования с характерной морфологией Х. четко различимы в световом микроскопе лишь в период клеточного деления - в метафазе и ( рис. 1 , 2 ). Основу Х. на всех стадиях клеточного цикла составляют -нитевидные структуры, которые во время деления клетки плотно закручены, обусловливая ,а в неделящейся клетке раскручены (деспирализованы). При завершении деления клетки разошедшиеся к её полюсам Х. разрыхляются и окружаются ядерной мембраной. В период между двумя делениями клетки (эта стадия клеточного цикла называется ) деспирализация Х. продолжается и они становятся малодоступными для наблюдения в световой микроскоп. Морфология Х. существенно отличается от таковой у и вирусов. Прокариоты (доядерные) и вирусы содержат обычно одну линейную или кольцевую Х., которая не имеет надмолекулярной укладки и не отделена от цитоплазмы ядерной оболочкой. Понятие Х. к генетическому аппарату прокариот применимо лишь условно, т.к. оно сформировалось при изучении Х. эукариот и подразумевает наличие в Х. не только сложного комплекса биополимеров ( и белков), но и специфической надмолекулярной структуры. Поэтому ниже даётся описание только Х. эукариот. Изменения внешнего вида Х. в клеточном и жизненном циклах обусловлены особенностями функционирования Х. Общий же принцип их организации, индивидуальность и непрерывность Х. в ряду клеточных поколений и организмов сохраняются неизменными. Доказательства тому получены при биохимическом, цитологическом и генетическом исследованиях Х. разных организмов. Они легли в основу . Молекулярные основы строения Х. Значение Х. как клеточных органоидов, ответственных за хранение, воспроизведение и реализацию наследственной информации, определяется свойствами биополимеров, входящих в их состав. Первая молекулярная модель Х. была предложена в 1928 Н. К. ,предугадавшим принципы их организации. Запись наследственной информации в Х. обеспечивается строением молекулы (ДНК), её .В Х. сосредоточено около 99% всей ДНК клетки, остальная часть ДНК находится в других клеточных органоидах, определяя цитоплазматическую наследственность. ДНК в Х. эукариот находится в комплексе с основными белками - и с негистоновыми белками, которые обеспечивают сложную упаковку ДНК в Х. и регуляцию её способности к синтезу (РНК) - .
Х. в интерфазе. Х. выполняет свои основные функции - репродукцию и транскрипцию - в интерфазе, поэтому строение Х. на этой стадии клеточного цикла представляет особый интерес. В интерфазе Х. плохо различимы потому, что в связи с активным синтезом РНК многие участки Х. (т. н.
) сильно раскручены; другие же (
) не участвуют в синтезе РНК и продолжают сохранять плотную упаковку (см. также
)
.В эухроматиновых участках, помимо элементарных дезоксирибонуклеопротеидных нитей (ДНП), имеются рибонуклеопротеидные частицы диаметром 200-500
Для изучения интерфазных Х. используют либо биохимические методы выделения вещества Х. - и разделения его на эухроматин и гетерохроматин, либо электронно-микроскопическое исследование интактных ядер и изолированного хроматина; как модели интерфазных Х. используют гигантские Х. типа ламповых щёток из ооцитов животных и многонитчатые (политенные) Х. двукрылых. В Х. типа ламповых щёток неактивные участки имеют вид плотно упакованных структур - ( рис. 2 , 3 ), которые обнаруживаются и в Х. соматических клеток, особенно в профазе митоза, и рассматриваются как характерные морфологические, а возможно и функциональные, единицы Х. В участках Х., активно синтезирующих РНК, хромомеры раскручиваются и образуют боковые петли, в которых молекулы РНК, соединяясь с белком, образуют рибонуклеопротеиды (РНП) - частицы, представляющие собой форму упаковки генных продуктов и различающиеся в отдельных боковых петлях по размерам и морфологическим признакам. В политенных Х., возникающих в тканях двукрылых и некоторых растений за счёт многократной репликации (удвоения) исходной Х. без последующего расхождения дочерних Х. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 |
|||||||
 , называемые РНП-гранулами, интергранулами и перихроматиновыми гранулами. Эти частицы представляют собой форму упаковки РНК, синтезированной на Х. и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму.
, называемые РНП-гранулами, интергранулами и перихроматиновыми гранулами. Эти частицы представляют собой форму упаковки РНК, синтезированной на Х. и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму.