Основы зоопсихологии
ModernLib.Net / Фабри Курт / Основы зоопсихологии - Чтение
(стр. 12)
|
Автор:
|
Фабри Курт |
|
Жанр:
|
|
|
-
Читать книгу полностью
(814 Кб)
- Скачать в формате fb2
(802 Кб)
- Скачать в формате doc
(279 Кб)
- Скачать в формате txt
(272 Кб)
- Скачать в формате html
(827 Кб)
- Страницы:
1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28
|
|
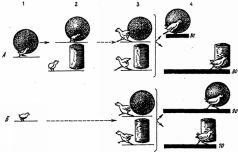
Как показали эксперименты, проведенные Фабри на цыплятах, первоначальный эффект врожденного узнавания при реакции следования недолговечен и без постоянного упражнения довольно быстро угасает. Однако этот эффект, т.е. следование, можно легко восстановить путем факультативного научения, направленного на объект запечатления. В упомянутых экспериментах объектом запечатления служил для одних цыплят синий шар, для других - желтый цилиндр. После первых же предъявлений цыплята стали следовать за этими объектами не менее интенсивно и продолжительно, чем в обычных условиях за курицей. Но даже наиболее активные подопытные цыплята переставали следовать за объектом запечатления уже через несколько дней, если прекращались ежедневные тренировки. В обычных же условиях, как известно, эта реакция сохраняется в течение ряда недель. Однако и в искусственных условиях эксперимента цыплята тем дольше продолжали следовать за объектом запечатления, чем больше было всего пробежек, т.е. длительность сохранения реакции следования оказалась прямо пропорциональной интенсивности локомоторной активности (частоте пробежек). В дальнейшем, уже после того, как реакция следования у цыплят совсем исчезла, у них в специальной установке были выработаны навыки следования за бывшим объектом следования - синим шаром или соответственно желтым цилиндром. Следование за этим объектом подкреплялось пищей. Другими словами, следование было заново выработано у цыплят, но путем факультативного научения. Результаты этой части эксперимента показали, что у цыплят, у которых раньше была сформирована реакция следования путем запечатления, этот навык образовался намного быстрее, чем у цыплят, которые до этого вообще не подвергались запечатлению, т.е. не имели возможности следовать за каким-либо подвижным объектом или же следовали за объектом запечатления другой группы подопытных животных (для цыплят, запечатленных на синий шар, это был желтый цилиндр и наоборот, - рис. 21). Из этих данных, казалось бы, следует, что врожденное действие (само следование), протекание которого не зависит от побочных внешних факторов и сроки проявления которого генетически фиксированы, угасает без упражнения так же, как это происходит с навыками, а с другой стороны, навык, факультативное научение, может вновь «оживить» эту инстинктивную реакцию. Объясняется это, очевидно, тем, что существует не одна, а две генетически различные, но функционально тождественные формы следования - истинно врожденная и индивидуально приобретаемая, причем вторая форма образуется путем чисто факультативного научения (ведь курица подкрепляет следование своих цыплят кормом и обогревом). Очевидно также, что первая форма со временем все больше перерастает во вторую, иными словами, реакция следования претерпевает качественную перестройку, теряя свою первоначальную врожденную сущность. Одновременно врожденное узнавание переходит в благоприобретаемое узнавание, основанное на индивидуальном опыте. Рис. 21. Формирование следования за подвижным объектом путем запечатления и факультативного научения (опыты
Фабри). А -экспериментальная группа цыплят, Б - контрольная группа цыплят.
1- выработка и тренировка реакции следования за синим шаром путем запечатления (1-4-е сутки после вылупления); 2 - первый контрольный опыт; цыплята следуют за объектом запечатления (синим шаром), но не за другим подвижным объектом (желтым цилиндром) (5-е сутки); 3 - второй контрольный опыт (26-е сутки), проведенный после трехнедельного перерыва, во время которого цыплята находились в обычных условиях, но не имели возможности видеть шар и цилиндр. В результате цыплята не только не следуют за этими движущимися объектами, но даже пугаются их. Так же реагируют на движущиеся шар и цилиндр контрольные (не подвергшиеся запечатлению) цыплята; 4 - выработка навыка следования за двигающимися шаром или желтым цилиндром при одновременном их предъявлении (27-31-е сутки) путем факультативного научения. Столбики с цифрами показывают среднее число сочетаний, необходимых для формирования устойчивого следования
Все это указывает на значение раннего факультативного (а не только облигатного) научения для полноценного проявления инстинктивных элементов поведения. Иными словами, уже на первых этапах постнатального развития врожденное поведение нуждается в участии научения в обеих его формах, которые при этом сливаются с ним. Этим прежде всего и характеризуется сущность всего поведения животных на ранних этапах постнатального онтогенеза.
Раннее формирование общения
Раннее формирование общения у птиц
Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда, коммуникативные компоненты поведения уже неоднократно встречались нами при ознакомлении с разными аспектами онтогенеза поведения, особенно в процессах запечатления, одним из определяющих факторов которого и является общение.
У птиц, как было уже показано (в частности, в предыдущей главе на кайрах), установление акустических контактов между родительской особью и ее потомством начинается уже на эмбриональной стадии развития птенцов путем пренатального облигатного научения (эмбрионального запечатления). У кайр, например, этот процесс происходит в форме установления акустико-кинестетических связей. В результате этого запечатления птенец подбегает к источнику запечатленного звука. Эта реакция обнаруживается даже у только что вылупившегося птенца: услышав запечатленный звук, он подбегает к родительской особи (в эксперименте к замещающему ее громкоговорителю), издает ответный крик и залезает под нее (или под аппарат). Если же из громкоговорителя после этого раздается другой звук (чужой зов), то птенец немедленно выскакивает из-под него и прячется в укрытии (опыты Чанц). Таким образом, коммуникативный компонент врожденного пускового механизма достраивается в данном случае в ходе эмбрионального запечатления.
Вместе
стем и у родительской особи происходит запечатление индивидуальных особенностей голоса еще не вылупившегося птенца, что обеспечивает индивидуальное опознавание последнего уже к моменту вылупления. Подобное «обратное запечатление» было установлено и у других птиц, а в постнатальном периоде также у млекопитающих и ряда рыб.
Укажем еще на работы Готтлиба, который показал на утятах, что если эмбрион не подвергается в последние дни инкубации соответствующим акустическим воздействиям, то установление звуковых контактов между вылупившимся птенцом и материнской особью будет значительно затруднено. Изучая затем реакцию следования у нескольких видов уток (крякв, пекинских уток, мандаринок), Готтлиб установил, что наиболее активно утята следовали за макетом (чучелом) при видотипичной визуально-акустической стимуляции, хуже - при одной лишь акустической, т.е. без чучела, и еще хуже при одной оптической, т.е. при неозвученном чучеле. Вместе с тем реакция следования значительно ослабевала, когда видотипичные визуальные или акустические компоненты (позывные звуки и внешний вид чучела) подменялись невидотипичными (т.е. принадлежащими другому виду уток). Канадский ученый Р.М.Эванс в опытах над чайками показал, что искажения в реакциях вылупившихся птенцов (предпочитание криков другого вида) могут обусловливаться отсутствием соответствующего эмбрионального опыта, а именно когда эмбрион лишен возможности слышать крики птиц своего вида.
Итак, как правило, вылупившиеся птенцы реагируют четко избирательно на видотипичные крики, т.е. предпочитают ориентироваться по «натуральным» акустическим раздражителям с большой биологической валентностью. Эти реакции основаны на врожденном узнавании (и предпочтении) видотипичных компонентов коммуникации, которое дополняется и уточняется облигатным выучиванием индивидуальных компонентов общения в процессе пре- и особенно постнатального запечатления. Следовательно, при постнатальном формировании акустического общения между птенцами и их родителями возможность индивидуального (не врожденного!) опознавания особей обеспечивается ранним облигатным научением. Принадлежность же последних к одному и тому же виду устанавливается птенцами на основе врожденного узнавания.
На основе данных, полученных рядом ученых, сейчас можно, очевидно, считать установленным, что и другие функции пения птиц формируются в большой степени благодаря постнатальному облигатному научению. Например, у овсянки песня молодого самца приобретает сексуальную, а затем и территориальную функцию только после того, как он научается петь «полную» песню, т.е. модифицировать ювенильную «подпесню», придать ей специфическую структуру и видоизменить ритм.
Большую роль играет при этом подражание, на что, в частности, еще в середине 50-х годов указывал Торп, отмечая, что птицы учатся непосредственно по звуковым сигналам других птиц.
Выше уже рассматривались явления подражания и говорилось о необходимости выделить как отдельные категории облигатное и факультативное имитационное научение. Теперь должно быть ясно, что облигатное подражание, будучи особым проявлением облигатного научения, характерно прежде всего для раннего постнатального периода онтогенеза. В области звукового общения это можно хорошо проиллюстрировать примерами из многочисленных полевых экспериментов, осуществленных советскими орнитологами К.А. и К.К.Вилкс путем перемещения яиц и птенцов в гнезда других видов (еще раньше подобные эксперименты проводились Промптовым). Изучив поведение приблизительно 4000 пересаженных воробьиных птиц (преимущественно мухоловок-пеструшек), эти исследователи установили, что птенцы в исключительно широком диапазоне перенимают песни «приемных родителей»: более 80% самцов, выросших в гнездах других видов, имитировали песню вида, к которому принадлежала особь, воспитавшая птенца (горихвостки, пеночки-трещотки, большой синицы). Вместе с тем при имитации чужой песни воспроизводились и элементы песни собственного вида. Понятно, что в нормальных условиях акустическое поведение птенца формируется путем облигатного имитационного научения звукам, издаваемым кормящей птицей того же вида.
Примечательно, что перенимание чужих звуков происходит в относительно короткий период начиная с вылета птенца из гнезда. Это доказывается тем, что чужие звуки имитировали и птицы, пересаженные незадолго перед вылетом, но если, наоборот, птенец воспитывался с самого начала в чужом гнезде, но перед вылетом пересаживался обратно к своему виду, чужое воспитание не сказывалось на акустических реакциях такой птицы и ее пение оставалось видотипичным. Таким образом, здесь определенно можно говорить о наличии сенсибильного периода, что опять-таки доказывает, что мы имеем в этом случае дело с облигатным научением, точнее, облигатным имитационным научением.
Раннее формирование
общения млекопитающих
У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет - в зависимости от степени зрелорождения, но чаще всего в первые часы жизни новорожденного. Детеныш одногорбого верблюда, например, издает первые звуки еще во время самих родов, а уже час спустя он в состоянии воспроизвести почти все звуки, свойственные его виду, поэтому уже с момента рождения начинается интенсивный акустический контакт между ним и его матерью.
Американским исследователем онтогенеза поведения животных Дж. П. Скоттом и рядом других ученых было доказано, что у собак и других видов псовых существует сенсибильный период, в течение которого только и возможно такое установление контактов с материнской особью и остальными щенками данного выводка. Очевидно, при этом процесс запечатления имеет комплексный характер, поскольку включает в себя и половое запечатление. На это указывает и то обстоятельство, что общение между матерью и щенками, равно как между последними и другими представителями того же вида, может быть по форме идентично половому общению (М. В. Фокс). Так же, по свидетельству ряда ученых, обстоит дело и у обезьян.
Большое значение для формирования общения имеет положительная тигмотаксисная реакция детенышей, выражающаяся в активном установлении физического контакта с поверхностью матери и собратьев, в прижимании к ним. Об этом свидетельствуют, в частности, эксперименты, проведенные на детенышах обезьян Г. Харлоу и его сотрудниками, применявшими разного рода макеты материнских особей (проволочные, тряпочные, обогревамые, холодные и пр.). Было показано, что детеныши предпочитают запечатленный холодный макет даже обогреваемому. Сходная тигмотаксисная реакция, как показали английский этолог В. Слэкин и другие исследователи, присуща также цыплятам и другим птенцам.
Скотт пришел к выводу, что у млекопитающих в раннем постнатальном онтогенезе существуют сенсибильные периоды трех типов: для процессов научения, стимуляции физиологических процессов и формирования общения. Исследования многих ученых показали, что если в течение сенсибильного периода последнего типа детеныш не имеет возможности установить связи с особями своего вида, он впоследствии может оказаться совершенно неспособным к общению с себе подобными и во всяком случае будет испытывать большие затруднения в коммуникативном, а также воспроизводительном поведении. То, что формирование первичных коммуникативных связей совершается путем запечатления, подтверждается и тем, что оно, как доказано рядом экспериментаторов, не зависит от пищевого или иного внешнего подкрепления.
Не менее четко выражено у млекопитающих и «обратное запечатление» - запечатление индивидуальных признаков детенышей у родителей. Так, например, овцы, козы и другие копытные научаются распознавать эти признаки непосредственно после рождения их детенышей и после этого уже не подпускают к соскам чужих. Ведущее значение имеет здесь, вероятно, ольфакторное опознавание, т.е. запоминание индивидуального запаха потомства, хотя и оптические признаки могут играть существенную роль. У детеныша, как показали в эксперименте с чернохвостыми оленями Д. и Х.Мюллер-Шварце, также происходит запечатление индивидуального запаха матери или, в эксперименте, заменяющей модели, причем в отношении последней вполне осуществляются акты общения, проявляемые в норме по отношению к подлинной матери.
Формирование общения у низших позвоночных
Отмеченные закономерности не имеют конечно, значения всеобщих правил для всех позвоночных, а тем более беспозвоночных. Даже среди птиц и млекопитающих обнаруживаются многочисленные отклонения и исключения, обусловленные экологическими факторами, особенностями образа жизни. С другой стороны, эти закономерности проявляются и у многих рыб. Так, например, мальки цихлид также научаются опознавать своих родителей путем запечатления и неотступно следуют за ними, но только тогда, когда те находятся в брачном наряде. Таким образом, облигатное научение здесь более крепко связано с врожденным узнаванием и более ограничено непосредственным действием ключевых раздражителей (в данном случае светло-темные контрасты, зеленые и синие цвета, движение), чем это наблюдается у высших позвоночных. Доминирующую роль здесь играют врожденные пусковые механизмы, индивидуальное же распознавание играет второстепенную роль. Средствами общения являются при этом сами движения, выполняемые родителями и мальками и имеющие генетически фиксированное сигнальное значение.
Что же касается «обратного запечатления», то оно у цихлид не связано с индивидуальным опознаванием потомства родительскими особями. Последние узнают мальков лишь как таковых, но неспособны отличить свой выводок от чужого, как это показали опыты с перекрестным воспитанием молоди путем замены родителей в семейных группах. При этом более крупная молодь поедается. Если подрастающих мальков повторно заменить более молодыми (т.е. более мелкими), то уход за потомством со стороны родителей можно растянуть на срок до 9 месяцев. На этой фазе преобладает оптическое общение, на личиночной - химическое (опыты Д.Наукса и Дж.Барлоу).
У низших позвоночных, в частности у рыб, присутствие особей того же вида не является столь необходимым для формирования форм общения, как у высших животных. В этом также проявляется более жесткое врожденное «программирование» раннего постнатального поведения рыб, чем у последних. Сходное положение наблюдается, очевидно, и у пресмыкающихся. Так, у ящериц-анолисов важный компонент общения - кивание головой с одновременным вытягиванием ног - наблюдается уже у новорожденных (через 5 минут после вылупления), причем и у изолированных животных, лишенных возможности видеть сородичей. Таким образом, здесь налицо подлинные врожденные двигательные координации, жестко регламентирующие коммуникативное поведение без индивидуального опознавания сородичей.
Познавательные аспекты раннего постнатального поведения
Исследовательское поведение
Постоянной составной частью любого поведенческого акта является исследовательское поведение, проявляющееся на разных онто- и филогенетических уровнях в весьма различных формах. Диапазон этих проявлений простирается от элементарных ориентировочных реакций до исследовательской деятельности высших млекопитающих.
В широком смысле ориентировочное, поисковое поведение является начальной фазой любого инстинктивного действия, и оно направлено на поиск тех агентов среды, которые являются объектами врожденного узнавания. Иначе говоря, в инстинктивных поведенческих актах ориентировочно-исследовательская активность служит поиску и обнаружению ключевых раздражителей и направляет на них деятельность животного. Об этом уже говорилось при изложении этологической концепции инстинктивного поведения.
Элементарные ориентировочные реакции проявляются очень рано даже у незрелорождающихся детенышей. Так, у щенят и лисят уже в первый-второй день жизни наблюдаются поисковые маятникообразные движения головой, которые прекращаются после нахождения соска матери. Сходные поисковые движения, повороты головы в стороны и тыканье концом морды в окружающие животное объекты производятся в первые же сутки жизни также котятами и многими другими детенышами хищных млекопитающих. Детеныши обезьян уже в первые дни жизни двигают ушными раковинами и т.д.
В дальнейшем появляются такие ориентировочные реакции, как обнюхивание, прислушивание, присматривание и т.п. У щенят и лисят такое обследование окружающего пространства начинается приблизительное конце второй недели жизни. У детенышей низших обезьян начиная с 14-16-го дня жизни появляются двигательные реакции в ответ на «индифферентные» звуковые и зрительные раздражители, которые потом (приблизительно в 1,5-2-месячном возрасте) вырастают в подлинные исследовательские действия: животное живо реагирует на изменения в окружающей среде (появление нового раздражителя, нового объекта) поворотами головы, особенно глаз, движениями ушных раковин, а также приближением к новому, незнакомому объекту. Так появляется «любопытство», столь характерное прежде всего для приматов, а также для других высших позвоночных.
Развитие исследовательского поведения в большой мере определяется условиями, в которых растет детеныш, особенно возможностями общения с матерью и другими сородичами. В экспериментах с длительным (до 30 недель) изолированным выращиванием детенышей макаков-резусов (опыты американских ученых Ч.М.Байзингера, Е.М.Брандта и Дж.Мичелля) было установлено, что изолированные детеныши больше рассматривали окружающие их предметы, человека, наблюдавшего за ними, и самих себя, чем нормально живущие со своими матерями. Первые также больше манипулировали предметами и больше трогали самих себя, вторые больше, быстрее и ловчее передвигались по клетке. Чехословацкие исследователи Б.Семигиновский, З.Халупка и И.Мысливечек, ставя опыты над крысятами, нашли, что возможность усиленного общения с сородичами в течение первых 6 недель жизни несколько тормозит развитие исследовательской активности, а обогащенная среда, наоборот, стимулирует его.
Уже на начальных стадиях постнатального онтогенеза наряду с реакциями на появление новых раздражителей имеет место активный поиск последних (маятникообразные движения головой у щенков), который особенно характерен для исследовательского поведения. Лежащие в основе этого поведения ориентировочные рефлексы отличаются, как показали исследования Е.Н.Соколова, значительной сложностью их физиологических механизмов. По Соколову, ориентировочные реакции возникают в результате несовпадения воспринимаемых новых сигналов с формирующимися в головном мозгу «нервными моделями стимула». При этом происходит оценка нового стимула, побуждающая животное к выполнению определенных действий или к их изменению. Наряду с этим ориентировочная деятельность играет также существенную роль в обеспечении необходимого уровня физиологической активности центральной нервной системы.
Нас же в данном случае особенно интересует познавательное значение ориентировочно-исследовательской деятельности, ее роль в приобретении новой информации о компонентах окружающей среды, в обогащении индивидуального опыта животного.
Облигатное научение и ориентация
Рассмотрим сначала некоторые процессы, связанные с начальной ориентацией у детеныша. У всех животных здесь первостепенное значение имеют таксисы, которые, как уже было показано, у высших животных дополняются и обогащаются элементами научения.
Это научение носит прежде всего характер облигатного научения, ибо, как только что отмечалось, оно входит как обязательный компонент в любой инстинктивный акт. Однако каждое животное вынуждено самостоятельно выучивать значимые лишь для него ориентиры, которые будут для разных особей того же вида весьма различными. Отличительные признаки этих ориентиров являются сами по себе случайными, несущественными, и только индивидуальное запоминание их в результате факультативного научения придает им опознавательное значение. Уже говорилось о том, что их выбор животным определяется в большой степени индивидуальными особенностями последнего и разные особи могут предпочитать разные компоненты среды в качестве ориентиров.
Таким образом, в ориентировочном поведении детеныша всегда присутствуют элементы облигатного и факультативного научения, однако соотношения между этими двумя компонентами, их удельный вес могут быть различными в зависимости от того, в какой функциональной сфере совершается ориентация. Когда детеныш учится распознавать «живые ориентиры», особенно внешние признаки своих сородичей, т.е. когда ориентация сочетается с общением, явно преобладают моменты облигатного научения. Это вполне понятно, ибо когда животное ориентируется по подаваемым другими животными оптическим, акустическим, ольфакторным и иным сигналам, которые входят в репертуар общения, то решающее значение имеют наследственно фиксированные, врожденные элементы поведения. При формировании коммуникативного поведения в онтогенезе первостепенное значение имеет поэтому срочная постнатальная достройка соответствующих врожденных пусковых механизмов, что, как мы уже знаем, является главной характерной чертой запечатления. Ограничимся здесь отсылкой и к другим аспектам и примерам облигатного научения с ориентационным значением (например, привыкание к «биологически нейтральным» раздражителям), о которых речь шла выше.
Раннее факультативное научение и ориентация
Уже в раннем ориентировочном поведении заметно сказываются индивидуальные особенности животного. В большой степени индивидуальные различия в поведении зависят от частоты и характера осуществленных с момента рождения сенсорных действий. Это достаточно четко проявляется при выращивании детенышей в условиях, когда им приходится постоянно видеть определенные фигуры. Как показали экспериментальные исследования, животные, выросшие в таком окружении, впоследствии легче ориентируются по таким фигурам. Правда, согласно экспериментам американского ученого Р.М.Осволта, существенное значение имеет при этом степень трудности различения таких фигур. В этих экспериментах крысята еще до прозрения помещались в клетки, на стенах которых попарно помещались фигуры: для одних это были горизонтальные и вертикальные полосы, для других - дуги, для третьих - треугольники и круги. Заранее было известно, что последнее сочетание является наиболее трудным для различения этими животными, полосы же - наиболее легким. В таком окружении крысята пребывали до 50-дневного возраста. Контрольные животные выращивались в клетках без фигур (с белыми поверхностями). Когда затем перед крысятами была поставлена задача - произвести выбор между знакомыми фигурами (одна из которых подкреплялась пищей, другая - нет), то оказалось, что ранний сенсорный опыт облегчал решение задачи только тогда, когда животным приходилось ориентироваться по трудноразличимым фигурам (треугольник и круг). Выращивание же с легче различимыми фигурами не давало такого эффекта: соответственные подопытные животные ориентировались по полосам и дугам не лучше контрольных. При этом Осволт отмечает, что влияние «преэкспозиции» стимулов явно приурочено к определенному раннему периоду, что указывает на участие в данном процессе наряду с факультативными также облигатных компонентов латентного научения.
Итак, ранняя факультативная неподкрепляемая визуальная тренировка способствует ориентации при дальнейшем подкрепляемом факультативном научении (заучивании ориентиров), особенно тогда, когда эта тренировка была сопряжена с трудностями и если имели место более сложные сенсорные действия.
В целом приведенные факты ранней пространственной ориентации показывают, как облигатное научение (благо приобретаемая ориентация по биологически валентным элементам среды) сочетается с факультативным научением, дополняется и конкретизируется им (запоминание особенностей местности путем сенсомоторной тренировки и заучивание конкретных ориентиров). При пространственной ориентации описанного типа на первый план выступает факультативный компонент. Иное соотношение наблюдается, как мы видели, в тех случаях, когда животное ориентируется не по признакам местности, а по признакам сородичей или других животных.
Раннее манипулирование
Исключительно большое значение имеет для приобретения и обогащения индивидуального опыта, как и для всей познавательной деятельности животного, манипулирование. Под манипулированием, манипуляционной активностью мы понимаем активное обращение с различными предметами при преимущественном участии передних, реже - задних конечностей, а также других эффекторов: челюстного аппарата, хобота (у слона), хватательного хвоста (у широконосых обезьян), щупалец (у головоногих моллюсков), клешней (у раков) и т.д. Манипулирование проявляется прежде всего в пищедобывательной и гнездостроительной активности животных. Вообще же, за исключением некоторых ориентировочных, сигнальных и защитных движений, при которых меняется лишь внешний вид животного, но само оно остается на месте, все движения, направленные на внешнюю среду, т.е. составляющие поведение, подразделяются на локомоторные (у большинства высших животных - опорно-локомоторные) и манипуляционные.
У высших животных, как еще будет показано, манипулирование выступает как ведущий фактор развития сенсомоторных функций. Это обусловлено тем, что именно в ходе манипулирования животное вступает в наиболее активный контакт с предметными компонентами среды и получает наилучшие возможности для ознакомления с ними, а также для многообразного воздействия на них.
Манипулирование появляется в различные сроки и проявляется по-разному у разных животных. Особенно велики эти различия у зрело- и незрелорождающихся видов. Но в каждом случае, выполняя разнообразные действия с различными предметами (биологически значимыми и «нейтральными»), детеныши получают комплексную информацию об объекте манипулирования, его свойствах, особенно о его физической структуре. Одновременно происходит развитие и совершенствование (путем тренировки) эффекторносенсорных систем животного. Все это делает манипуляционную активность высшей формой ориентировочно-исследовательской деятельности животных.
В полной мере манипулирование развертывается в игровом периоде онтогенеза. В рассматриваемом здесь периоде манипуляционная активность появляется первоначально лишь в самых простых формах, особенно у незрелорождающихся животных. Так, например, детеныши хищных млекопитающих в первые дни или даже недели, а именно до начала функционирования дистантных рецепторов, т.е. до открывания глаз и ушных проходов, в основном спят, а в промежутках преимущественно ползают в поисках соска и сосут.
По данным советского этолога Н.Н.Мешковой, первые движения детенышей носят характер простейшего манипулирования: детеныши трогают лапой родителей и своих собратьев, хватают их ртом в разных местах тела, нажимают на них мордочкой. Все эти действия еще весьма нечеткие и непродолжительные. У лисенка, по исследованиям того же автора, до 12-дневного возраста (до прозрения) манипуляционная (нелокомоторная) активность развивается следующим образом.
Первые манипуляционные движения обнаруживаются уже у новорожденного, но в течение первых двух часов после рождения они существуют лишь в двух формах: 1) прикасания к объекту передним концом головы (конкретно это проявляется в поиске соска) и 2) хватания объекта губами (захват соска и прилегающих участков кожи ртом). Передние конечности в этих действиях участия не принимают и отведены в стороны.
Затем в течение первых двух суток с момента рождения к этим двум формам добавляются еще пять, которые на этом этапе также связаны только с сосанием и сопутствующими ему действиями (расталкивание других детенышей при движении к соску, поиск соска, отгребание шерсти на животе матери, придавливание или ритмичное толкание лапами живота рядом с соском во время сосания). Эти манипуляции характеризуются прежде всего тем, что детеныш начинает действовать передними лапами и появляются боковые движения головы (в частности, при расталкивании собратьев). Передние лапы прикасаются к объекту (телу матери) и передвигаются по нему одновременно с головой или же самостоятельно, без ее участия.
Последнее имеет место в тех случаях, когда лисенок разгребает одними передними лапами шерсть на животе матери или придавливает его сразу обеими лапами.
Страницы:
1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28
|
|
