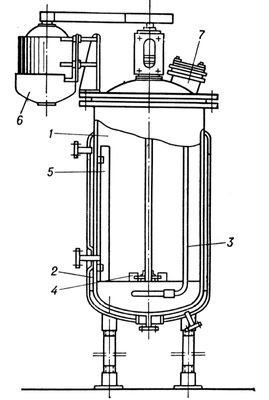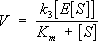
,
где
k
3и
К
т–константы, характерные для каждой реакции.
Эта зависимость, установленная экспериментально для многих ферментативных реакций, может быть теоретически выведена, если превращение субстрата в продукт реакции (Р) происходит по механизму образования и распада комплекса между ферментом и субстратом –
ES-комплекса:
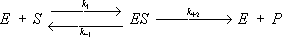
,
где
k
1, k
-1и
k
+ 2–константы, характеризующие скорость указанных стрелками стадий процесса, причём соотношение (
k
-1+ k
+ 2)
/ k
-1= К
т.Если в реакции участвует не один, а несколько (в большинстве случаев два) субстратов и
ES-комплекс образует продукты реакции не в одну, а в несколько стадий, зависимость выражается более сложными уравнениями, однако и они могут быть выведены лишь на основе представления о первичном образовании
ES-комплексов. Для многих ферментов получены прямые доказательства образования
ES-комплексов. Так, спектральными методами доказано образование комплексов с участием дегидрогеназ и пероксидаз; выделены в кристаллич. состоянии комплексы оксидазы
D-аминокислот с
D-aланином, карбоксипептидазы А с глицил-
L-тирозином. В ряде случаев установлено пространственное строение
ES-комплексов методом рентгеноструктурного анализа.
Высокая специфичность Ф. к. объясняется строгим геометрическим и электронным соответствием структуры субстрата структуре
фермента, на котором субстрат сорбируется и далее претерпевает химические превращения. Допускается, что соответствие (комплементарность) геометрического и электронного строения активного центра и реагирующих с ним участков молекулы субстрата (субстратов) достигается в момент сближения субстрата с активным центром (гипотеза индуцированного соответствия Д. Э. Кошленда, США). Активный центр фермента, представляющий собой ансамбль химически активных группировок (функциональных групп аминокислот), формируется из остатков аминокислот, нередко расположенных далеко друг от друга в полипептидной цепи, но сближенных в пространстве в результате глобулярной структуры белка. Часто в построении активных центров участвуют низкомолекулярные вещества (ионы металлов, органические кофакторы). В молекуле a-химотрипсина, катализирующего гидролиз белков и полипептидов и имеющего цепь длиной в 246 аминокислотных остатков, активный центр образован остатками серина (порядковый номер остатка в цепи 195), гистидина (№ 57), изолейцина (№ 16) и аспарагиновой кислоты (№ 102 и № 194). Активный центр рибонуклеазы, катализирующей расщепление РНК и построенной из 124 аминокислот, образован остатками лизина (№ 7 и № 41), аргинина (№ 39) и гистидина (№ 12 и № 119). Активные центры мн. ферментов функционируют с участием низкомолекулярных веществ –
Ф. к. К ним относятся производные витаминов,
,а также ионы некоторых металлов (Na, К, Ca, Mg, Zn, Fe, Сu, Со, Mo и др.).
Общая теория Ф. к. не разработана, однако результаты исследования механизма действия ферментов позволяют качественно, а в отдельных случаях и количественно объяснить высокую активность Ф. к. Её главные причины: 1) сближение реагентов при сорбции их в активном центре, этот фактор эквивалентен повышению концентрации реагирующих веществ; 2) специфическая ориентация сорбированного в активном центре субстрата, благоприятная для взаимодействия с каталитическим участком активного центра; 3) образование химических связей между субстратом и каталитическим участком активного центра, направляющее реакцию по энергетически наиболее лёгкому пути; 4) осуществление всех основных химических превращений субстрата «внутримолекулярно» – в составе фермент-субстратного комплекса; 5) исключительная гибкость молекулы фермента, позволяющая активному центру принимать на каждой стадии превращения фермент-субстратного комплекса строение, способствующее достижению максимальной скорости данной стадии реакции. Каждая предшествующая стадия подготавливает наилучшие условия для последующей. Ориентировочная оценка суммарного эффекта всех перечисленных факторов Ф. к. позволяет теоретически предсказать возможное ускорение реакции в 10
10–10
13раз, что во многих случаях совпадает с найденной экспериментально величиной.
Механизмы регуляции активности Ф. к. связаны с особенностями белковой структуры ферментов. Глобулярное строение ферментов, поддерживаемое относительно слабыми химическими связями между отдельными участками полипептидной цепи, легко нарушается при изменении кислотности среды, температуры, концентрации солей в клетках и т.п. Поскольку для Ф. к. необходима строго заданная структура фермента, все эти факторы оказывают воздействие на его активность. Каждый фермент максимально активен при определённой температуре, pH среды и т.п. Изменение условий среды в обе стороны от оптимума снижает активность Ф. к.; нередко она саморегулируется продуктом реакции. Для обратимых процессов, когда фермент катализирует прямую и обратную реакции, скорость прямой реакции (активность Ф. к.) уменьшается при образовании избытка продукта реакции.
Важную роль в Ф. к. играет т. н. аллостерическая регуляция активности ферментов. В живой клетке совершается множество последовательных химических реакций, катализируемых соответствующими ферментами
E
1
,
E
2и т.п.
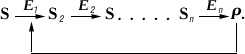
Обнаружены многочисленные реакции, когда образующийся в избытке против физиологически необходимых количеств продукт
Рспособен снижать активность первого фермента
E
1и тем самым уменьшать скорость всей цепи реакций. Такой механизм называется регуляцией по принципу обратной связи. При этом регулятор Р (в общем случае носит наименование эффектор) воздействует на специальный регуляторный центр фермента
E
1
,расположенный вдали от активного центра. Однако вследствие подвижности структуры белковой молекулы фермента в целом реакция с регуляторным центром приводит к изменению строения и свойств активного центра. Такой участок получил, по предложению Ф.
и Ж.
,наименование аллостерического центра, а сами ферменты типа
E
1называется аллостерическими ферментами. В качестве аллостерических эффекторов часто выступают нуклеотиды (например, адениловая кислота, аденозинтрифосфат и т.п.) и аминокислоты (в реакциях биосинтеза др. аминокислот).
К аллостерическим относят также механизмы регуляции действия фермента, содержащего несколько активных центров, при которых связывание субстрата в активном центре вызывает изменение (уменьшение или увеличение) активности фермента. Аллостерическими свойствами обладают ферменты, построенные из нескольких (чётного числа) молекул, каждая из которых имеет активный и регуляторный центры. Воздействие эффектора на регуляторный центр одной из молекул вызывает общее (кооперативное) изменение строения в др. молекулах и активности фермента в целом. Возможны также регуляторные механизмы, при которых воздействие эффектора на аллостерический фермент приводит к изменению степени ассоциации составляющих его субъединиц, что также сопровождается изменением общей активности фермента. Такого рода механизмы играют важную роль в регуляции сложной системы химических реакций (
) в живом организме.
Лит.:«Журнал Всес. химического общества им. Д. И. Менделеева», 1971, т. 16, № 4; Дженке В, П., Катализ в химии и энзимологии, пер. с англ., М., 1972: Структура и функции активных центров ферментов. Сб., посвященный 70-летию со дня рождения А. Е. Браунштейна, М., 1974.
В. А. Яковлев.
Ферментёр
Ферментёр,аппарат для глубинного выращивания (культивирования) микроорганизмов в питательной среде в условиях стерильности, интенсивного перемешивания, непрерывного продувания стерильным воздухом и постоянной температуры. Ф. представляет собой герметичный цилиндрический сосуд – корпус, снабженный барботером для подачи стерильного воздуха и мешалкой с электроприводом. Внутри Ф. вдоль его корпуса и перпендикулярно к нему закрепляют узкие металлические полосы – отбойники для повышения эффективности перемешивания. Объём Ф., предназначенных для лабораторных исследований, чаще до 30
л, для полузаводских экспериментов – 0,05–5
м
3
,промышленного использования – 50–100
м
3
.Лабораторные Ф. могут изготовляться из термостойкого стекла (их стерилизуют в автоклавах), Ф. больших размеров – из нержавеющей стали (они имеют паровую рубашку для стерилизации и поддержания температуры). Ф., как правило, оборудуются устройствами для измерения и регулирования температуры, количества продуваемого воздуха и давления внутри Ф. В случае необходимости Ф. дополнительно снабжается устройствами для измерения и регулирования pH среды, концентрации растворённого кислорода в культуральной жидкости, углекислого газа в выходящем воздухе, сигнализатором уровня пены и приспособлениями для механического или химического пеногашения. При непрерывном процессе культивирования микроорганизмов Ф. дополнительно оборудуются стерилизуемыми резервуарами для хранения компонентов питательной среды и насосами для их непрерывной подачи в Ф. Используют Ф. в промышленности при
антибиотиков, ферментов, витаминов, аминокислот, нуклеотидов, белково-витаминных концентратов и т.д., в научных исследованиях в области микробиологии, биохимии и др. родственных дисциплин.
Лит.:Уэбб Ф., Биохимическая технология и микробиологический синтез, пер. с англ., М., 1969; Производство антибиотиков, М., 1970.
М. А. Гильзин.
Ферментёр: 1 — корпус; 2 — паровая рубашка; 3 — барботёр; 4 — мешалка; 5 — отбойник; 6 — электропривод; 7 — загрузочный люк.
«Ферментная и спиртовая промышленность»
«Ферме'нтная и спиртова'я промы'шленность»,научно-технический и производственный журнал, орган министерства пищевой промышленности СССР и центрального правления научно-технического общества пищевой промышленности. Периодичность 8 номеров в год. Издаётся в Москве с 1924: название менялось (в частности, с 1953 по 1963 назывался «Спиртовая промышленность»). Освещает достижения науки и техники в спиртовой, пивобезалкогольной, ликёро-водочной, ферментной и ацетонобутиловой промышленности, опыт передовых предприятий. Тираж (1975) 4600 экз.
Ферментные препараты
Ферме'нтные препара'ты,лекарственные средства, содержащие
,оказывают направленное влияние на обмен веществ. Ф. п. получают из продуктов животного происхождения, растений и микроорганизмов. Желудочный сок,
,
и др. Ф. п. и ферменты применяют при желудочно-кишечных заболеваниях с нарушением функций желёз органов пищеварения. Широкое применение в медицинской практике нашли Ф. п. протеолитического действия (см.
)
,получаемые из поджелудочной железы крупного рогатого скота (например,
)
.Они расщепляют пептидные связи в белках и пептидах. Трипсин при местном воздействии разрушает некротизированные ткани и фибринозные образования, разжижает вязкие секреты, экссудат, сгустки крови, при внутримышечном введении оказывает противовоспалительное действие. Применяют трипсин в виде ингаляций или внутримышечно для облегчения удаления секрета и экссудата при бронхитах, бронхоэктатической болезни; при лечении тромбофлебита, остеомиелита, гайморита, иридоциклита и др. заболеваний; местно – при лечении ожогов, пролежней, гнойных ран. Дезоксирибонуклеаза уменьшает вязкость гноя, задерживает развитие вирусов герпеса, аденовирусов; применяют при герпетических и аденовирусных заболеваниях глаз, абсцессах лёгких, поражениях верхних дыхательных путей. Препарат лидаза, содержащий фермент гиалуронидазу, вызывает увеличение проницаемости тканей и облегчает движение жидкостей в межтканевых пространствах; применяют при контрактурах суставов, рубцах после ожогов и операций, гематомах и др. Для лечения тромбоэмболий, тромбофлебитов, инфаркта миокарда применяют фибринолизин, растворяющий свежие тромбы. Пенициллиназа, получаемая из культуры Bacillus cereus, инактивирует препараты пенициллина, в связи с чем применяется при аллергических реакциях, вызванных этими препаратами.
В медицинской практике применяют также препараты с антиферментной активностью:
(угнетают холинэстеразу), некоторые антидепрессивные средства (угнетают моноаминоксидазу); в качестве мочегонных – ингибиторы карбоангидразы (например, диакарб); при острых панкреатитах – ингибиторы протеолитических ферментов (например, трасилол).
Лит.:Капланский С. Я., Применение ферментных препаратов в терапии различных заболеваний, в кн.: Актуальные вопросы современной биохимии, т. 2, М., 1962; Машковский М. Д., Лекарственные средства, 7 изд., ч. 2, М., 1972.
В. В. Чурюканов.
Ферментные яды
Ферме'нтные я'ды,вещества различной химической природы, специфически подавляющие активность определённого
или группы родственных ферментов. По существу Ф. я. представляют собой
ферментов, которые даже в очень низких концентрациях угнетают жизненно важные физиологические функции организма. Многие ядовитые вещества, т. н. «нервные яды» (люизит), «дыхательные яды» (цианиды, H
2S), пестициды (ядохимикаты) оказывают отравляющее действие в результате ингибирования отдельных ферментов (например, холинэстеразы у членистоногих). Изучение влияния Ф. я. на изолированные ферменты или ферментные системы позволяет целенаправленно искать эффективные противоядия к определённым отравляющим веществам или новые пестициды для борьбы с вредными насекомыми, клещами и т.д. и сорняками. Иногда термин «Ф. я.» применяют для обозначения ферментов, входящих в состав ядов змей, пчёл, скорпионов и др. и разрушающих клетки крови или др. тканей человека и животных.
Ферментопатии
Ферментопа'тии,энзимопатии, заболевания, обусловленные врождённым дефектом обмена веществ вследствие ферментных нарушений; относятся к группе
.В основе Ф. лежат различные виды нарушений (полное отсутствие фермента, снижение его активности, отсутствие или неправильный синтез кофермента и др.), последствия которых в виде определённых аномалий обмена веществ и определяют в каждом случае специфику клинической картины Ф. Например, аномалии углеводного обмена могут проявляться в виде сахарного диабета, галактоземии; жирового обмена – в виде болезней Тей-Сакса, Нимана-Пика; аминокислотного обмена – в виде алкаптонурии,
и т.п. Известно около 500 видов Ф. Многие из них отличаются полиморфизмом и т. н. гетерогенностью, которая заключается в том, что аномалии различных генов, регулирующих взаимодействие ферментов, могут иметь идентичные проявления, т.к. ферменты, контролирующие разные биохимические реакции, нередко дают одинаковый конечный результат метаболизма. Большинство Ф. передаётся по аутосомно-рецессивному типу наследования. Некоторые Ф. могут быть выявлены с помощью экспресс-методов в первые дни жизни ребёнка, например
.Во многих случаях ранняя диагностика Ф. позволяет нормализовать обмен веществ с помощью специально подобранной диеты, введения в организм недостающего вещества (заместительная терапия), гормонов или удаления избытка продуктов метаболизма, нарушающего обмен веществ. Перспективен также метод внутриутробной диагностики (изучение культивируемых клеток околоплодной жидкости, реже – прямое исследование её). В профилактике Ф. возрастает роль
(см. также
,
)
.
Лит.:Бадалян Л. О., Таболин В. А., Вельтищев Ю. Е., Наследственные болезни у детей, М., 1971; Харрис Г., Основы биохимической генетики человека, пер. с англ., М., 1973; Howell R. R., Moore Ch. М., Prenatal diagnosis in the prevention of genetic disease, «Texas medicine», 1974, v. 70, № 5, p. 77–84.
Ферменты
Ферме'нты(от лат. fermentum – закваска), энзимы, специфические белковые катализаторы, присутствующие во всех живых клетках. Почти все биохимические реакции, протекающие в любом организме и в своём закономерном сочетании составляющие его
,катализируются соответствующими Ф. Направляя и регулируя обмен веществ, Ф. играют важнейшую роль во всех процессах жизнедеятельности.
Как всякие
,Ф. снижают
,необходимую для осуществления той или иной химической реакции, направляя её обходным путём – через промежуточные реакции, которые требуют значительно меньшей энергии активации. Так, реакция АБ ® А + Б в присутствии Ф. идёт следующим образом: АБ + Ф ® АБФ и далее АБФ ® БФ + А и БФ ® Б + Ф. Например, для осуществления реакции гидролиза дисахарида сахарозы, в результате которого образуются глюкоза и фруктоза, без участия катализатора требуется 32 000
кал(1
кал= 4,19
дж) на моль сахарозы. Если же реакция катализируется Ф. b-фруктофуранозидазой, то необходимая энергия активации составляет всего 9400
кал.Подобное понижение энергии активации под влиянием Ф. – следствие перераспределения электронных плотностей и некоторой деформации молекул субстрата, происходящей при образовании промежуточного соединения – фермент-субстратного комплекса (АБФ). Эта деформация, ослабляя внутримолекулярные связи, приводит к понижению необходимой энергии активации и, следовательно, ускоряет течение реакции (см.
,
)
.
История изучения ферментов.В 1814 рус. химик К. Г. С.
открыл ферментативное действие водных вытяжек из проросшего ячменя, расщеплявших крахмал до сахара. Можно считать, что эти работы положили начало энзимологии (ферментологии) как самостоятельному разделу биологической химии. В 1833 французскими химиками А. Пайеном и Ж. Персо впервые был выделен из солода препарат фермента амилазы, что способствовало развитию препаративной химии Ф. В середины 19 в. разгорелась дискуссия о природе
между Л.
,с одной стороны, и Ю.
,П. Э. М.
и К.
–с другой. Опираясь на свои классические работы, Пастер развивал представление о том, что брожение вызывается лишь живыми микроорганизмами и что процесс брожения неразрывно связан с их жизнедеятельностью. Либих и его сторонники, отстаивая химическую природу брожения, считали, что оно является следствием образования в клетках микроорганизмов растворимых Ф., подобных выделяемой из солода амилазе. Однако все попытки выделить из разрушенных дрожжевых клеток растворимый Ф., способный вызвать брожение, не удавались. Дискуссия Либиха и Пастера о природе брожения была разрешена в 1897 Э.
,который, растирая дрожжи с инфузорной землёй, выделил из них бесклеточный растворимый ферментный препарат (названный им зимазой), вызывавший спиртовое брожение. Открытие Бухнера утвердило материалистическое понимание природы брожений и имело большое значение для дальнейшего развития как энзимологии, так и всей биохимии.
В начале 20 в. Р.
с сотрудниками стал широко применять для выделения и очистки Ф. метод адсорбции (впервые предложен А. Я.
для разделения Ф. поджелудочной железы). Работы Вильштеттера, имевшие большое значение для характеристики свойств отдельных Ф., привели вместе с тем к принципиально неправильному выводу, что Ф. не принадлежат ни к одному из известных классов органических соединений. Выдающимся успехом в выяснении химической природы Ф. были исследования американских биохимиков Дж.
,выделившего в 1926 в кристаллическом виде Ф.
из семян канавалии, и Дж.
,получившего в 1930 кристаллы протеолитического Ф. пепсина. Работы Самнера и Нортропа указали путь получения высокоочищенных кристаллических препаратов Ф. и вместе с тем неопровержимо доказали белковую природу Ф.
С середине 20 в. благодаря развитию методов физико-химического анализа (главным образом
) и методов белковой химии расшифрована первичная структура многих Ф. Так, работами американских биохимиков С. Мура, У. Стайна и К. Анфинсена показано, что Ф. рибонуклеаза из поджелудочной железы быка представляет собой полипептидную цепочку, состоящую из 124 аминокислотных остатков, соединённых в 4 местах дисульфидными связями.
С помощью рентгеноструктурного анализа расшифрована вторичная и третичная структура ряда Ф. Так, методом рентгеноструктурного анализа английский учёный Д. Филлипс в 1965 установил трёхмерную структуру Ф.
.Показано, что многие Ф. обладают также четвертичной структурой, т. е. их молекула состоит из нескольких идентичных или различных по составу и структуре белковых субъединиц (см.
)
.
Общая характеристика ферментов.Все Ф. разделяются на две большие группы: однокомпонентные, состоящие исключительно из белка, и двухкомпонентные, состоящие из белка, называемого апоферментом, и небелковой части, называемой простетической группой. Апофермент двухкомпонентных Ф. называют также белковым носителем, а простетическую группу – активной группой. Благодаря работам О.
,А.
,Ф.
,Ф.
и Л. Лелуара установлено, что простетические группы многих Ф. представляют собой производные
или
.Т. о. была открыта важнейшая функциональная связь между Ф., витаминами и нуклеотидами, являющимися строительными «кирпичиками» нуклеиновых кислот.
Примером двухкомпонентного Ф. является
,катализирующая расщепление пировиноградной кислоты на двуокись углерода и уксусный альдегид: CH
3COCOOH ® CH
3CHO + CO
2. Простетическая группа пируватдекарбоксилазы (тиаминнирофосфат) образована молекулой тиамина (витамина B
1) и двумя остатками фосфорной кислоты. Простетические группы ряда важных окислительно-восстановительных Ф. – дегидрогеназ содержат производное амида никотиновой кислоты (ниацина), или же рибофлавина (витамина B
2); в состав простетических группы т. н.
,катализирующих перенос аминогрупп (–NH
2) и декарбоксилирование и ряд др. превращений аминокислот, входит пиридоксальфосфат – производное витамина B
6; активная группа Ф., катализирующих перенос остатков различных органических кислот (например, ацетила CH
3CO–), включает витамин
.К двухкомпонентным Ф. относятся также важные окислительные Ф. –
(катализирует реакцию разложения перекиси водорода на воду и кислород) и
(окисляет перекисями различные соединения, например полифенолы с образованием соответствующего хинона и воды). Каталитическое действие этих Ф. может быть воспроизведено с помощью ионов трёхвалентного железа. Эти ионы обладают, однако, очень малой каталитической активностью, которая может быть усилена, если атом железа входит в состав
.Хотя гем обладает уже значительным каталазным действием, его каталитическая активность всё же в несколько миллионов раз меньше активности каталазы, в которой гем в качестве простетической группы этого Ф. связан со специфическ им белком. Гем обладает также слабым пероксидазным действием, однако это действие проявляется в полной мере только после соединения гема со специфическим белком в целостный Ф. пероксидазу. Т. о., соединение простетической группы с белком приводит к резкому возрастанию её каталитической активности. Вместе с тем от природы белка зависит не только каталитическая активность, но и специфичность действия Ф. Прочность связи простетической группы и апофермента различна у разных Ф. У некоторых Ф., например у дегидрогеназ, катализирующих окисление различных субстратов путём отщепления водорода, эта связь является непрочной. Такие Ф. легко диссоциируют (например, при
)
и распадаются на простетическую группу и апофермент. Простетические группы, легко отделяющиеся от белковой части Ф., называются
.
Многие Ф. содержат металлы, без которых Ф. не активен. Эти металлы называются кофакторам и. Так, пероксидаза и каталаза содержат железо, аскорбинатоксидаза, катализирующая окисление аскорбиновой кислоты, – медь, алкогольдегидрогеназа, окисляющая спирты в соответствующие альдегиды, – цинк.
Специфичность и механизм действия ферментов.Действие Ф., в отличие от неорганических катализаторов, строго специфично и зависит от строения субстрата, на который Ф. действует. Прекрасным примером такой зависимости служит катализируемая аргиназой реакция гидролитического расщепления аминокислоты аргинина на орнитин и мочевину:
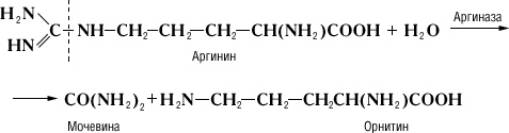
Однако аргиназа не расщепляет метилового эфира аргинина:
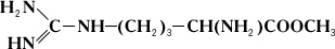
Дипептид, состоящий из остатков двух молекул аргинина, под действием аргиназы даёт лишь половину теоретического количества мочевины. Очевидно, что, хотя расщепление аргинина происходит в месте, весьма отдалённом от карбоксильной (COOH) группы (показано пунктиром), необходимым условием действия аргиназы является её соединение с карбоксильной группой аргинина. Поэтому замещение водорода в карбоксильной группе на метильный остаток или же связывание карбоксильной группы со второй молекулой аргинина оказывают резкое влияние на действие аргиназы. Примеры специфичности действия Ф. могут быть приведены при рассмотрении их стереохимической специфичности, т. е. действия Ф. на стереоизомеры (см.
)
.Так, Ф., окисляющий природные L-аминокислоты, не действует на D-изомеры этих же аминокислот; Ф. дипептидаза, гидролизирующий дипептиды, состоящие из остатков L-аминокислот, не действует на такие же дипептиды, состоящие из остатков D-аминокислот. Специфичность действия Ф. послужила нем. учёному Э.
основанием для сравнения субстрата и Ф., который катализирует его превращение, с замком и соответствующим ему ключом. Стереохимическая специфичность Ф. теснейшим образом связана с одной из основных особенностей живых организмов – их способностью к синтезу оптически активных органических соединений.
В образовании соединения между ферментом и субстратом – т. н. фермент-субстратного комплекса – принимают участие лишь некоторые функциональные группы молекулы Ф., образующие его
.Так, например, в молекуле гидролизирующего белки
,состоящего из 246 аминокислотных остатков, активный центр образован одним из остатков
(химотрипсин относится к сериновым протеиназам) и двумя остатками гистидина, расположенными в удалённых друг от друга участках полипептидной цепи. Сближение этих функциональных групп активного центра происходит благодаря свойственной молекуле химотрипсина специфической пространственной (третичной) структуре. Её нарушение в результате денатурации белка или каких-либо химических модификаций приводит к изменению или полной потере каталитической активности. В случае двухкомпонентных Ф. в образовании фермент-субстратного комплекса принимают участие не только функциональные группы апофермента, но и простетическая группа. Так, при расщеплении пировиноградной кислоты пируватдекарбоксилазой субстрат связывается с частью молекулы тиамин-пирофосфата следующим образом:
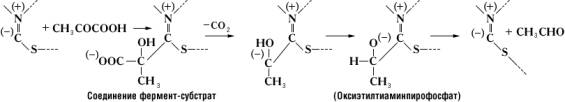
Исключительно высокая специфичность действия Ф. объясняется их белковой природой. Так, пиридоксалевые Ф., содержащие один и тот же кофермент (пиридоксальфосфат), могут принадлежать к различным классам и катализировать самые разнообразные реакции. Специфичность их действия зависит от природы апофермента.
Условия действия ферментов.Действие Ф. зависит от ряда факторов, прежде всего от температуры и реакции среды (pH). Оптимальная температура, при которой активность Ф. наиболее высока, находится обычно в пределах 40–50 °С. При более низких температурах скорость ферментативной реакции, как правило, снижается, а при температурах, близких к 0 °С, практически реакция полностью прекращается. При повышении температуры выше оптимальной скорость ферментативной реакции также снижается и, наконец, полностью прекращается. Снижение интенсивности действия Ф. при повышении температуры сверх оптимальной объясняется главным образом начинающимся разрушением (денатурацией) входящего в состав Ф. белка. Поскольку белки в сухом состоянии денатурируются значительно медленнее, чем белки оводнённые (в виде белкового геля или раствора), инактивирование Ф. в сухом состоянии происходит гораздо медленнее, чем в присутствии влаги. Поэтому сухие споры бактерий или сухие семена могут выдержать нагревание до гораздо более высоких температур, чем те же споры или семена в увлажнённом состоянии.
Важнейшим фактором, от которого зависит действие Ф., как установил впервые С.
,является активная реакция среды – pH. Отдельные Ф. различаются по оптимальной для их действия величине pH. Так, например, пепсин, содержащийся в желудочном соке, наиболее активен в сильнокислой среде (pH 1–2); трипсин – протеолитический Ф., выделяемый поджелудочной железой, имеет оптимум действия в слабощелочной среде (pH 8–9); оптимум действия папаина – протеолитического Ф. растительного происхождения – находится в слабокислой среде (pH 5–6).
Действие Ф. зависит также от присутствия специфических активаторов и неспецифических или специфических ингибиторов. Так, энтерокиназа, выделяемая поджелудочной железой, превращает неактивный трипсиноген в активный трипсин. Подобные неактивные Ф., содержащиеся в клетках и в секретах различных желёз, называются
.Многие Ф. активируются в присутствии соединений, содержащих сульфгидрильную группу (–SH). К ним принадлежат аминокислота цистеин и трипептид
,содержащийся в каждой живой клетке. Особенно сильное активирующее действие глутатион оказывает на некоторые протеолитические и окислительные Ф. Неспецифическое угнетение (ингибирование) Ф. происходит под действием различных веществ, дающих с белками нерастворимые осадки или блокирующих в них какие-либо группы (например, SH-группы). Существуют более специфические
Ф., угнетение которыми каталитических функций основано на специфическом связывании этих ингибиторов с определёнными химическими группировками в активном центре Ф. Так, окись углерода (CO) специфически ингибирует ряд окислительных Ф., содержащих в активном центре железо или медь. Вступая в химическое соединение с этими металлами, она блокирует активный центр Ф. и вследствие этого он теряет свою активность.