 |
|
Популярные авторы:: Лавкрафт Говард Филлипс :: Андерсон Пол Уильям :: Эллисон Харлан :: Ломер Кит :: Говард Роберт Ирвин :: Дойл Артур Конан :: Гилберт Честертон :: БСЭ :: Желязны Роджер :: Ламур Луис Популярные книги:: Справочник по реестру Windows XP :: Бурый волк :: Последний коммунист :: Тень в зеркале :: Дао - путь воды :: Смерть, какую ты заслужил :: Шотландский лев :: Снежная страна :: Злой гений Нью-Йорка [Дело Епископа] :: Лопатка |
Большая Советская Энциклопедия (ГЕ)ModernLib.Net / Энциклопедии / БСЭ / Большая Советская Энциклопедия (ГЕ) - Чтение (стр. 22)
признаков - «полимерными» генами, т. е. системами из многих генов. Многих признаки и болезни человека, наследующиеся сцепленно с полом, обусловлены генами, локализованными в
Х-или
Y-xpomocome. Таких генов известно около 120. К ним относятся гены гемофилии
Аи
В, недостаточности фермента глюкозо-6-фосфатдегидрогеназы, цветовой слепоты и др. Др. метод Г. ч. - близнецовый метод (см.
Близнецы
). Однояйцевые близнецы (ОБ) развиваются из одной яйцеклетки, оплодотворённой одним спермием; поэтому набор генов (
генотип
) у ОБ идентичен. Разнояйцевые близнецы (РБ) развиваются из двух и более яйцеклеток, оплодотворённых разными спермиями; поэтому их генотипы различаются так же, как у братьев и сестёр (сибсов). Сравнение внутрипарных различий между ОБ и РБ позволяет судить об относительном значении наследственности и среды в определении свойств человеческого организма. В близнецовых исследованиях особенно важен показатель конкордантности, выражающий (в %) вероятность обладания данным признаком одним из членов пары ОБ или РБ, если его имеет другой член пары. Если признак детерминирован преимущественно наследственными факторами, то процент конкордантности намного выше у ОБ, чем у РБ. Например, конкордантность по
группам крови
, которые детерминированы только генетически, у ОБ равна 100%. При шизофрении конкордантность у ОБ достигает 67%, в то время как у РБ - 12,1%; при врождённом слабоумии (олигофрении) - 94,5% и 42,6% соответственно. Подобные сравнения проведены в отношении ряда заболеваний. Т. о., исследования близнецов показывают, что вклад наследственности и среды в развитие самых разнообразных признаков различен и признаки развиваются в результате взаимодействия генотипа и внешней среды. Одни признаки обусловлены преимущественно генотипом, при формировании др. признаков генотип выступает в качестве предрасполагающего фактора (или фактора, лимитирующего норму реакции организма на действия внешней среды).
Геном человека включает несколько миллионов генов, способных к тому же по-разному влиять на развитие признаков. В результате мутаций и перекомбинации генов возникает присущее человеку разнообразие по самым разным признакам. Гены человека мутируют каждый с частотой от 1 на 100000 до 1 на 100000000 гамет на поколение. Распространение мутаций среди больших групп населения изучает популяционная Г. ч., позволяющая составить карты распространения генов, определяющих развитие нормальных признаков и наследственных болезней. Особый интерес для популяционной Г. ч. представляют изоляты - группы населения, в которых по каким-либо причинам (географическим, экономическим, социальным, религиозным и др.) браки заключаются чаще между членами группы. Это приводит к повышению частоты кровного родства вступающих в брак, а значит, и вероятности того, что рецессивные гены перейдут в гомозиготное состояние и проявятся, что особенно заметно при малочисленности изолята. Исследования в области Г. ч. продемонстрировали наличие естественного отбора в человеческих популяциях. Однако отбор у человека приобретает специфические черты: он интенсивно действует только на эмбриональной стадии (т. н. самопроизвольные аборты - отражение такого отбора). Отбор в человеческом обществе осуществляется посредством дифференциальной брачности и плодовитости, т. е. в результате взаимодействия социальных и биологических факторов. Мутационный процесс и отбор обусловливают огромное разнообразие (полиморфизм) по ряду признаков, присущее человеку, что делает его с биологической точки зрения необычайно пластичным и приспособленным видом. Широкое использование в Г. ч. цитологических методов способствовало развитию цитогенетики , где основной объект исследования - хромосомы , т. е. структуры клеточного ядра, в которых локализованы гены. Установлено (1946), что хромосомный набор в клетках тела человека (соматических) состоит из 46 хромосом, причём женский пол определяется наличием двух Х-хромосом, а мужской - Х-хромосомы и Y-xpomocomы. В зрелых половых клетках находится половинное (гаплоидное) число хромосом. Митоз , мейоз и оплодотворение поддерживают преемственность и постоянство хромосомного набора как в ряду клеточных поколений, так и в поколениях организмов. В результате нарушений указанных процессов могут возникать аномалии хромосомного набора с изменением числа и структуры хромосом, что приводит к возникновению т. н. хромосомных болезней, которые нередко выражаются в слабоумии, развитии тяжёлых врождённых уродств, аномалий половой дифференцировки или обусловливают самопроизвольные аборты. Успехи в развитии Г. ч. сделали возможными предупреждение и лечение наследственных заболеваний . Один из эффективных методов их предупреждения - медико-генетическое консультирование с предсказанием риска появления больного в потомстве лиц, страдающих данным заболеванием или имеющих больного родственника. Достижения биохимической Г. ч. раскрыли первопричину (молекулярный механизм) многих наследственно обусловленных дефектов, аномалий обмена веществ, что способствовало разработке методов экспресс-диагностики, позволяющих быстро и рано выявлять больных, и лечения многих прежде неизлечимых наследственных болезней. Чаще всего лечение состоит во введении в организм веществ, не образующихся в нём вследствие генетического дефекта, или в составлении специальных диет, из которых устранены вещества, оказывающие токсическое действие на организм в результате наследственно обусловленной неспособности к их расщеплению. Многие генетические дефекты исправляются с помощью своевременного хирургического. вмешательства или педагогической коррекции. Практические мероприятия, направленные на поддержание наследственного здоровья человека, на охрану генофонда человечества, осуществляются через систему медико-генетических консультаций . Основная цель медико-генетические консультирования - информировать заинтересованных лиц о вероятности риска появления в потомстве больных. К медико-генетическим мероприятиям относится также пропаганда генетических знаний среди населения, т.к. это способствует более ответственному подходу к деторождению. Медико-генетическая консультация воздерживается от мер принудительного или поощрительного характера в вопросах деторождения или вступления в брак, принимая на себя лишь функцию информации. Большое значение имеет система мер, направленных на создание наилучших условий для проявления положительных наследственных задатков и предотвращение вредных воздействий среды на наследственность человека. Г. ч. представляет собой естественнонаучную основу борьбы с расизмом , убедительно показывая, что расы - это формы адаптации человека к конкретным условиям среды (климатическим и иным), что они отличаются друг от друга не наличием «хороших» или «плохих» генов, а частотой распространения обычных генов, свойственных всем расам. Г. ч. показывает, что все расы равноценны (но не одинаковы) с биологической точки зрения и обладают равными возможностями для развития, определяемого не генетическими, а социально-историческими условиями. Констатация биологических наследственных различий между отдельными людьми или расами не может служить основанием для каких-либо выводов морального, юридического или социального порядка, ущемляющих права этих людей или рас (см. Геноцид , Сегрегация ). Лит.:Ниль Дж. и Шэлл У. Наследственность человека, пер с англ., М, 1958; Канаев И. И., Близнецы, М. - Л., 1959; Штерн К., Основы генетики человека, пер. с англ., М., 1965; Маккьюсик В., Генетика человека, пер. с англ., М., 1967; Биология человека, пер. с англ., М. 1968: Эфроимсон В. П., Введение в медицинскую генетику, 2 изд., М., 1968: Основы цитогенетики человека, [М., 1969]; Li Ching-chun, Human genetics, N. Y., 1961. К. Н. Гринберг, А. А. Прокофьева-Бельговская. Генетико-автоматические процессы Гене'тико-автомати'ческие проце'ссы, вероятностные процессы, определяющие изменение частоты разных аллелей в популяции. В больших, свободно скрещивающихся популяциях в отсутствие отбора и давления мутаций соотношение аллелей, независимо от их абсолютной исходной частоты, должно сохраняться во всех поколениях. Однако в реальных, ограниченных по численности популяциях частота генов не остаётся постоянной не только под давлением мутаций и отбора, но и в силу случайных отклонений. Детальный анализ Г.-а. п. был проведён советскими генетиками Н. П. Дубининым (1931), Н. П. Дубининым и Д. Д. Ромашовым (1932), английским - Р. Фишером (1931) и американским - С. Райтом (1931). Случайные колебания частоты аллелей популяции связаны с тем, что распределение аллелей между гаметами и комбинирование гамет в зиготе - вероятностные процессы. Г.-а. п. оказывают несистематический эффект, т.к. частота аллелей в разных поколениях может повышаться или понижаться. В малых популяциях или в популяциях, которые распадаются под действием изоляционных механизмов на отдельные подгруппы, может происходить чисто случайная стабилизация аллелей (гомозиготы) или их элиминация; в результате довольно быстро проявляются новые стабилизированные комбинации генов. Наиболее отчётливо Г.-а. п. проявляются при возникновении новых изолированных популяций. Например, в секте меннонитов (Ланкастер, штат Пенсильвания, США), насчитывающей около 8000 человек, значителен процент карликов с многопалостью (13% меннонитов гетерозиготны по гену, который в гомозиготном состоянии обусловливает появление таких карликов); это объясняется тем, что члены секты вступают в брак только между собой, а такая изоляция способствует появлению гомозиготных индивидуумов. В больших популяциях Г.-а. п. не могут обусловить такой стабилизации или элиминации аллелей, т.к. влияние этих процессов компенсируется за счёт разных факторов в последующих поколениях или в разных подразделениях популяции. Теория Г.-а. п. объяснила генетические последствия изоляции, судьбу рецессивных мутаций на уровнях малых концентраций и эволюцию популяций по нейтральным признакам. Г.-а. п. объясняют многие расовые различия человека, возникшие без действия отбора. Наряду с термином «Г.-а. п.» широко используется термин «дрейф генов», предложенный С. Райтом. Советский генетик С. С. Четвериков, подчёркивая роль вероятностно-статистических закономерностей при дрейфе генов, предлагал назвать это явление генетико-стохастическими процессами. Лит.:Дубинин Н. П., Эволюция популяции и радиация, М., 1966, с. 421-33. Н. П. Дубинин, В. Н. Сойер. Генетиков и селекционеров общество Гене'тиков и селекционе'ров о'бществоВсесоюзное имени Н. И. Вавилова (ВОГИС) научно-общественная организация при АН СССР, объединяющая ученых и практиков СССР, ра6отающих в области генетики и селекции . Создано в 1965. 30-31 мая 1966 Москве состоялся учредительный съезд общества; был утвержден устав общества избраны центральный совет (80 чел.) и президиум (23 чел.; находится в Москве). Президентом избран Б. Л. Астауров. К началу 1971 общество насчитывало 3670 член, имело 26 отделений в столицах союзных республик, краевых, областных центрах и др. городах. Цели и задачи ВОГИС: активное участие в развитии всех отраслей генетики и селекции, повышение квалификации членов общества и реализация их исследований, популяризация и пропаганда новейших теоретических и практических достижений в области генетики и селекции, содействие преподаванию генетики и селекции в средней и высшей школе. В. Ф. Мирек. Генетическая информация Генети'ческая информа'ция, заложенная в наследственных структурах организмов (в хромосомах, цитоплазме, клеточных органеллах), получаемая от предков в виде совокупности генов информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот ) и связанных с ними функциях. У многоклеточных форм при половом размножении Г. и. передается из поколения в поколение через посредство половых клеток - гамет , единственная функция которых - передача и хранение Г. и. У микроорганизмов и вирусов имеются особые типы передачи Г. и. (см. Сексдукция , Трансдукция , Трансформация ). Г. и. заключена преимущественно в хромосомах , где она зашифрована в определённой линейной последовательности нуклеотидов в молекулах дезоксирибонуклеиновой кислоты - ДНК (см. Генетический код ). Г. и. реализуется в ходе онтогенеза - развития особи - передачей Г. и. от гена к признаку. Все клетки организма возникают в результате делений единственной исходной клетки - зиготы - и потому имеют один и тот же набор генов - потенциально одну и ту же Г. и. Специфичность клеток разных тканей определяется тем, что в них активны разные гены, т. е. реализуется не вся Г. и., а только её часть, необходимая для функционирования данной ткани. Ю. С. Демин. Генетические карты хромосом Генети'ческие ка'рты хромосо'м, схемы относительного расположения сцепленных между собой наследственных факторов - генов . Г. к. х. отображают реально существующий линейный порядок размещения генов в хромосомах (см. Цитологические карты хромосом ) и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам ) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности . Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом (см. Гаплоид ). Американский генетик Т. Х. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов (см. Диплоид ) не абсолютное; в некоторых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответственными участками; этот процесс носит название перекреста, или кроссинговера . Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния - морганида - соответствует частоте кроссинговера в 1%. Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры . Составить Г. к. х. можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы - около 400 генов, распределенных в 10 группах сцепления ( рис. 1 ). У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления ( рис. 2 ). При переносе генетического материала из клетки-донора в клетку-реципиент, например при конъюгации , кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. некоторых бактериофагов . См. также Генетика , Мутация . Лит.:Лобашев М. Е., Генетика, 2 изд., Л., 1967; Медведев Н. Н., Практическая генетика, 2 изд., М., 1968; Актуальные вопросы современной генетики. Сб. ст., М., 1966; Жакоб Ф., Вольман Э., Пол и генетика бактерий, пер. с англ., М., 1962; Бензер С., Тонкая структура гена, в сборнике: Молекулярная генетика, пер. с англ., М., 1963; Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Рейвин А. У., Эволюция генетики, пер. с англ., М., 1967; Мюнтцинг А., Генетика, пер. с англ., 2 изд., М., 1967: Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967. В. С. Андреев. 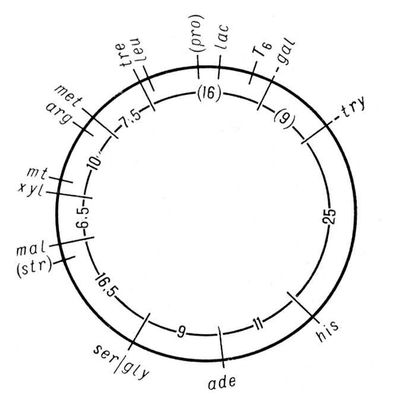
Рис. 2. Генетическая карта хромосомы кишечной палочки (Escherichia coli К 12). Цифры означают время (в мин), необходимое для переноса в клетку-реципиент генетических маркёров, контролирующих биосинтез ряда аминокислот, а также устойчивость к стрептомицину и к фагу Т 6; эти цифры характеризуют расстояние между генами. Обозначения: ade - аденин; his - гистидин; try - триптофан; gal - галактоза; lac - лактоза: pro - пролин; leu - лейцин; tre - треонин; met - метионин; arg - аргинин; mt - маннит; хуl - ксилоза; mal - мальтоза; ser - серин; gly - глицин; str и Т 6- устойчивость к стрептомицину или фагу T 6. 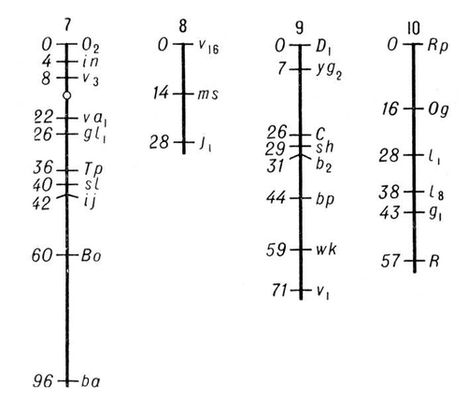
Рис. 1. Генетические карты 7-10 хромосом кукурузы. Цифры по длине хромосом обозначают расстояние от конца хромосомы в морганидах; буквы - сокращенные названия признаков, определяемых соответствующими генами. Генетические ряды Генети'ческие ряды', группы органических соединений с одинаковым числом атомов углерода в молекуле, различающихся функциональными группами; см. Гомологические ряды . Генетический анализ Генети'ческий ана'лиз, совокупность методов изучения наследственных свойств организмов. Г. а. включает: 1) Гибридологический метод, изучающий законы наследственности, а также строение и поведение наследственных структур с помощью специальных видов скрещиваний (см. Гибридологический анализ ). 2) Цитогенетический метод, развившийся на стыке генетики и цитологии. Главная его задача - установление связи между закономерностями наследования и строением и функциями хромосом (составление цитологических карт хромосом , геномный анализ и др.). 3) Молекулярно-генетический метод, получивший развитие в связи с новыми биохимическими и физико-химическими методами анализа наследственных структур. С его помощью изучается связь между молекулярным строением генов и синтезируемыми в соответствии с заложенной в них информацией белками . Лит.:Серебровский А. С.. Генетический анализ, М., 1970 (библ.). Ю. С. Демин. Генетический код Генети'ческий код, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов . В природных нуклеиновых кислотах - дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) - встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания , Пиримидиновые основания ). В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых кислот обнаружено около 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных сахаров. Т. к. количество кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. количество нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 4 2=16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Г. Гамов предложил (1954) модель триплетного Г. к., т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, наз. кодоном . Число возможных триплетов равно 4 3=64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (т. н. вырожденность кода). Было предложено много различных моделей Г. к., из которых серьёзного внимания заслуживали три модели (см. рис. ): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены следующие основные закономерности, касающиеся Г. к.: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Г. к.); 2) считывание Г. к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Г. к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Г. к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за некоторыми исключениями). Универсальность Г. к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (например, кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (например, проростков гороха), то в такой системе, будет идти белковый синтез. Благодаря работам американских генетиков М. Ниренберга, С. Очоа, Х. Корана известен не только состав, но и порядок нуклеотидов во всех кодонах (см. табл., построенную по данным опытов с кишечной палочкой). Из 64 кодонов у бактерий и фагов 3 кодона - УАА, УАГ и УГА - не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы , т. е. сигнализируют о завершении синтеза полипептида. Их назыают терминирующими кодонами. Существуют также 3 сигнала о начале синтеза - это т. н. инициирующие кодоны - АУГ, ГУГ и УУГ, - которые, будучи включенными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида. Реализация Г. к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит название транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК «переписывается» в нуклеотидную последовательность РНК. Второй этап - трансляция - протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке: этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки , раздел Биосинтез). Полный «словарь» генетического кода для аминокислот
* В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов. Лит.:Общая природа генетического кода для белков, в сборнике: Молекулярная генетика, пер. с англ., М., 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ., М., 1964, с. 9-23; Ниренберг М., Генетический код (II), там же, с. 24-41; Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер. с англ., М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М. - Л., 1966; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П.. Общая генетика, М., 1970. Н. П. Дубинин, В. Н. Сойфер. 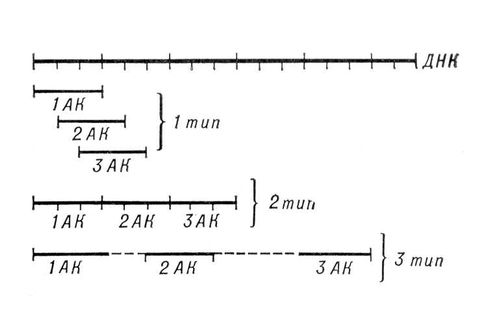
Модели генетического кода: 1-й тип - перекрывающийся код без запятых; 2-й тип - неперекрывающийся код без запятых; 3-й тип - код с «промежутками», т. е. код с запятыми. Генетический полиморфизм Генети'ческий полиморфи'зм, сосуществование в пределах популяции двух или нескольких различных наследственных форм, находящихся в динамическом равновесии в течение нескольких и даже многих поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (например, в разные сезоны), либо повышенной относительной жизнеспособностью гетерозигот . Один из видов Г. п. - сбалансированный Г. п. - характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от которого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают несколько форм Г. п., анализ которых позволяет определять действие отбора в природных популяциях. Лит.:Тимофеев-Ресовский Н. В., Свирежев Ю. М., О генетическом полиморфизме в популяциях, «Генетика», 1967, № 10. Генетическое действие излучений Генети'ческое де'йствие излуче'ний, радиационный мутагенез, возникновение наследственных изменений ( мутаций ) при облучении организмов. Г. д. и. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||